Explain each and everything about glycolysis
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
I now have everything needed for a comprehensive answer. Let me compile the full, detailed explanation of glycolysis.
Glycolysis - A Complete Explanation
What is Glycolysis?
Glycolysis (from Greek: glykys = sweet, lysis = splitting) is a universal metabolic pathway found in the cytosol of every living cell. It is the sequence of 10 enzymatic reactions that converts one molecule of glucose (6 carbons) into two molecules of pyruvate (3 carbons each), generating ATP and NADH in the process.
Glycolysis is truly at the hub of carbohydrate metabolism - virtually all sugars, whether from the diet or from internal catabolism, are eventually converted to glucose and funneled into this pathway. - Biochemistry, 8th ed, Lippincott Illustrated Reviews, p. 293
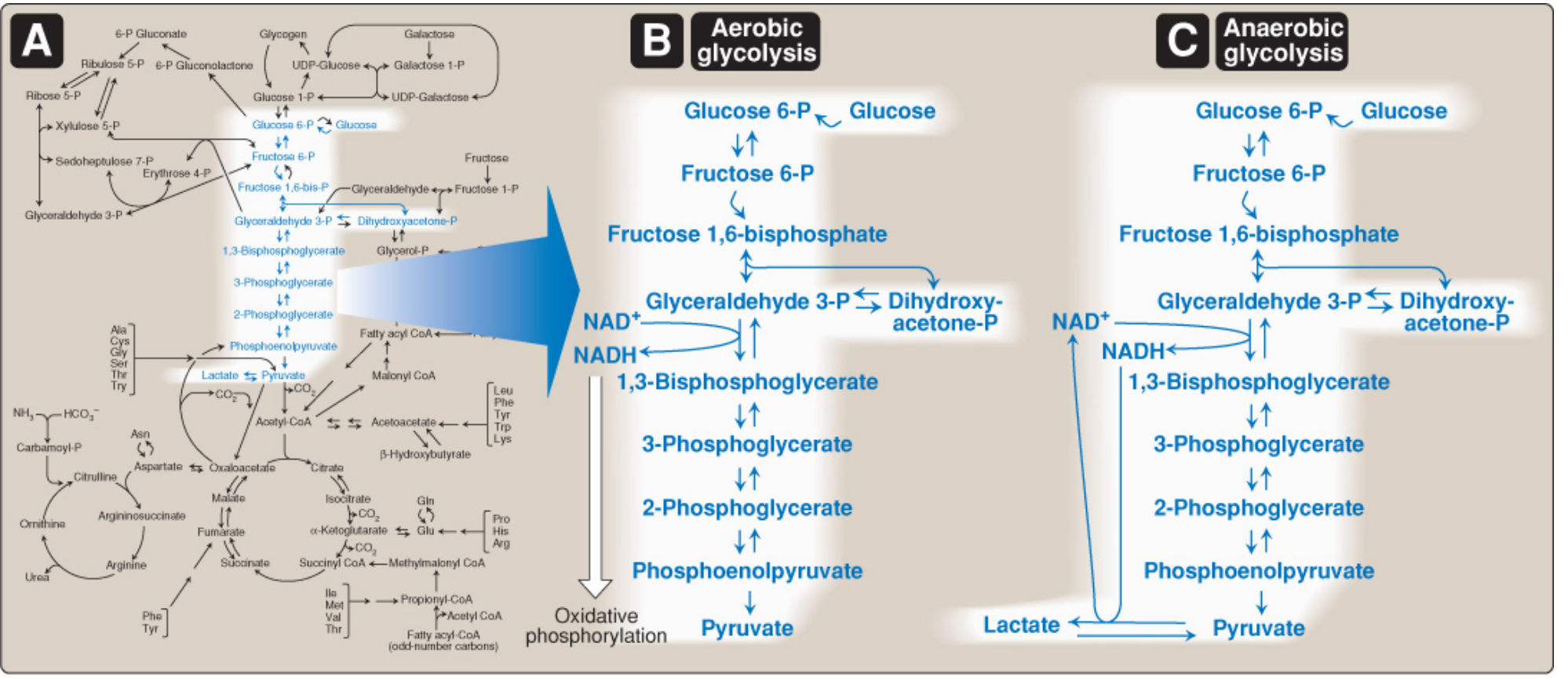
Figure: (A) Glycolysis as a hub of carbohydrate metabolism. (B) Aerobic glycolysis - pyruvate feeds oxidative phosphorylation. (C) Anaerobic glycolysis - pyruvate is reduced to lactate.
Location
Glycolysis occurs entirely in the cytosol (cytoplasm). No mitochondria are needed. This is why even cells without mitochondria (e.g., red blood cells) or cells deprived of oxygen can still perform glycolysis.
Overall Summary Reaction
Glucose + 2 NAD⁺ + 2 ADP + 2 Pi → 2 Pyruvate + 2 NADH + 2 H⁺ + 2 ATP + 2 H₂O
Net yield per glucose:
- 2 ATP (net; 4 produced, 2 consumed)
- 2 NADH
- 2 Pyruvate
The Two Phases
Glycolysis is divided into two distinct phases:
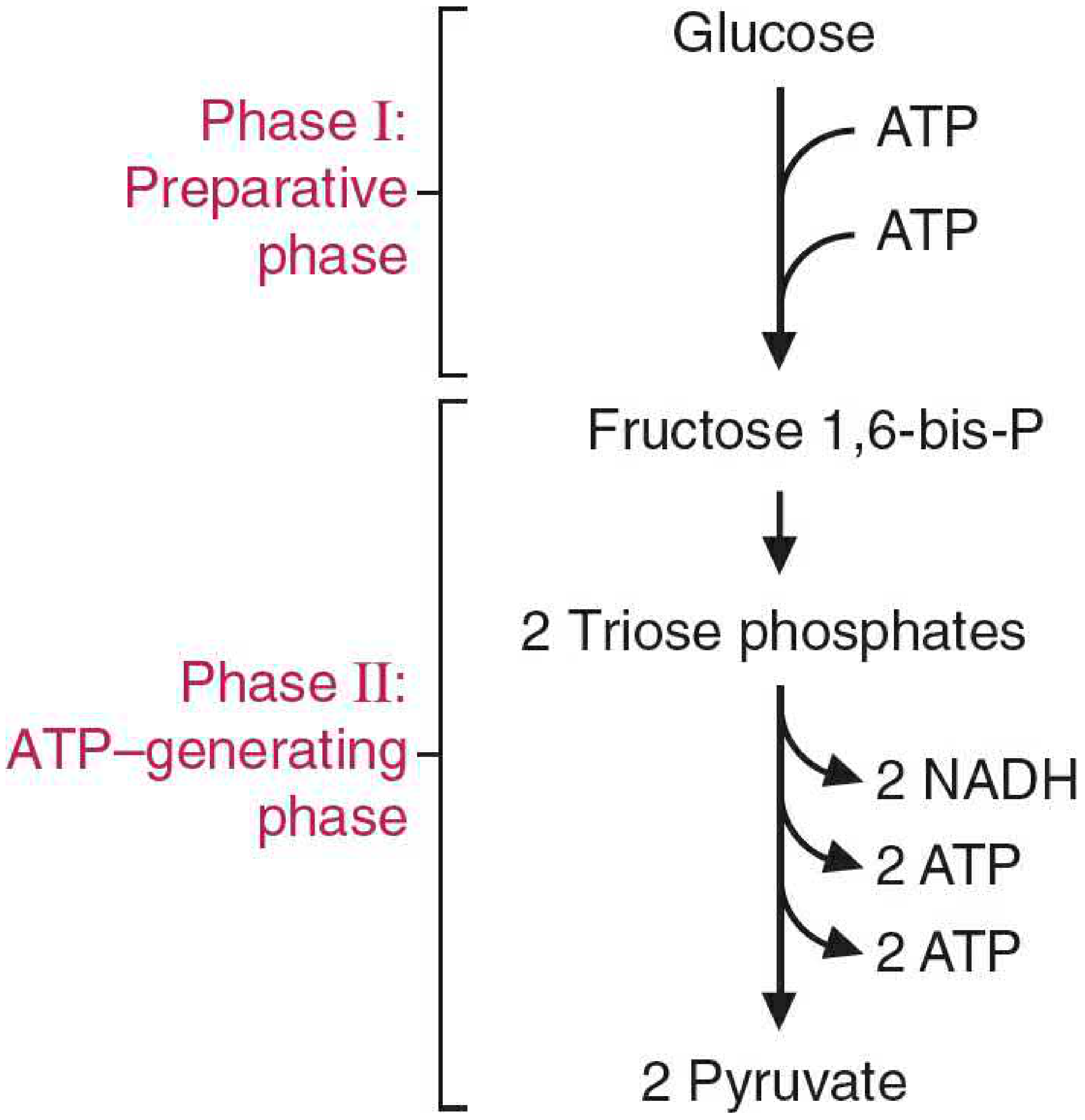
Phase 1: Energy-Investment (Preparatory) Phase - Steps 1-5
- Consumes 2 ATP to "prime the pump"
- Glucose is phosphorylated twice and split into two 3-carbon triose phosphates
- Goal: activate glucose for cleavage
Phase 2: Energy-Generation (Payoff) Phase - Steps 6-10
- Produces 4 ATP and 2 NADH
- The two triose phosphates are oxidized and dephosphorylated down to pyruvate
- Net gain: 2 ATP, 2 NADH
Glucose Transport Into Cells
Before glycolysis begins, glucose must enter the cell. It cannot diffuse freely across the lipid bilayer.
Two transport systems:
-
GLUT transporters (Na⁺/ATP-independent) - Passive facilitated diffusion via a family of 14 isoforms (GLUT-1 to GLUT-14). Each transports one molecule at a time (uniporter).
- GLUT-1: ubiquitous, most tissues
- GLUT-2: liver, kidneys, pancreatic β-cells (high Km, low-affinity, high-capacity - acts as glucose sensor)
- GLUT-4: skeletal muscle and adipose tissue (insulin-dependent - insulin increases GLUT-4 insertion into membrane)
- GLUT-5: fructose transporter in intestine
-
SGLT (Na⁺-dependent cotransport) - Active transport in intestinal epithelium and renal tubules; couples glucose uptake to the Na⁺ gradient.
The 10 Steps of Glycolysis in Detail
STEP 1: Glucose → Glucose 6-Phosphate (G6P)
Enzyme: Hexokinase (HK I-III in most tissues) / Glucokinase (HK IV in liver and pancreatic β-cells)
Cofactor: Mg²⁺-ATP
Reaction: Glucose + ATP → Glucose 6-phosphate + ADP
Energy: Consumes 1 ATP; reaction is irreversible (highly negative ΔG°)
Key points:
- Phosphorylation traps glucose inside the cell (charged phosphate group prevents membrane crossing)
- G6P is a branch-point metabolite - it can enter glycolysis, the pentose phosphate pathway, or glycogen synthesis
- Hexokinase (I-III): Low Km (~0.1 mM) = high affinity for glucose; functions at low glucose. Inhibited by its product G6P (product inhibition). Low Vmax - prevents overconsumption of phosphate
- Glucokinase (HK IV): High Km (~10 mM) = low affinity; only active during hyperglycemia (after a meal). High Vmax - rapidly processes a flood of glucose. NOT inhibited directly by G6P. In liver: regulated by glucokinase regulatory protein (GKRP) - sequestered in nucleus when fructose-6-P is high; released to cytosol when glucose rises. In pancreatic β-cells: acts as a glucose sensor for insulin secretion threshold.
STEP 2: Glucose 6-Phosphate → Fructose 6-Phosphate (F6P)
Enzyme: Phosphoglucose isomerase (PGI)
Reaction: G6P ⇌ F6P (aldose → ketose isomerization)
Energy: No ATP consumed or produced; reversible
Key points:
- Converts an aldose (glucose) to a ketose (fructose) sugar
- Positions the keto group next to carbon 3, which is essential for the next step (cleavage)
- Equilibrium favors G6P, but is pulled forward by consumption of F6P
STEP 3: Fructose 6-Phosphate → Fructose 1,6-Bisphosphate (F1,6-BP)
Enzyme: Phosphofructokinase-1 (PFK-1) - the most important regulatory enzyme of glycolysis
Cofactor: Mg²⁺-ATP
Reaction: F6P + ATP → Fructose 1,6-bisphosphate + ADP
Energy: Consumes 1 ATP; irreversible (committed step of glycolysis)
Key points:
- This is the rate-limiting, committed step - considered the main control point of glycolysis
- PFK-1 is an allosteric enzyme with a regulatory and catalytic site
- Activators of PFK-1: AMP, ADP, fructose 2,6-bisphosphate (F-2,6-BP), inorganic phosphate
- Inhibitors of PFK-1: ATP (at high concentrations), citrate (signals that TCA cycle intermediates are abundant)
- Fructose 2,6-bisphosphate (F-2,6-BP) is the most potent allosteric activator. It is synthesized by PFK-2 (stimulated by insulin) and degraded by fructose 2,6-bisphosphatase (stimulated by glucagon). This gives hormones indirect control over glycolytic flux.
STEP 4: Fructose 1,6-Bisphosphate → Glyceraldehyde 3-Phosphate + Dihydroxyacetone Phosphate
Enzyme: Aldolase
Reaction: F1,6-BP → Glyceraldehyde 3-phosphate (G3P) + Dihydroxyacetone phosphate (DHAP)
Energy: No ATP used or generated; reversible
Key points:
- The 6-carbon molecule is cleaved into two 3-carbon molecules
- Only G3P continues directly in glycolysis
- DHAP is isomerized to G3P in the next step (so effectively both halves proceed)
STEP 5: Dihydroxyacetone Phosphate → Glyceraldehyde 3-Phosphate
Enzyme: Triose phosphate isomerase (TPI)
Reaction: DHAP ⇌ Glyceraldehyde 3-phosphate
Energy: No ATP; reversible
Key points:
- Converts DHAP into a second G3P
- After this step, all further reactions proceed twice (once for each G3P from one glucose)
- TPI is one of the most catalytically efficient enzymes known (near "catalytic perfection")
From here on, every step happens TWICE per glucose molecule.
STEP 6: Glyceraldehyde 3-Phosphate → 1,3-Bisphosphoglycerate (1,3-BPG)
Enzyme: Glyceraldehyde 3-phosphate dehydrogenase (GAPDH)
Reaction: G3P + NAD⁺ + Pᵢ → 1,3-BPG + NADH + H⁺
Energy: No ATP consumed; 2 NADH produced (one per G3P = 2 total per glucose); uses inorganic phosphate
Key points:
- This is the only oxidation step in glycolysis - G3P (an aldehyde) is oxidized to a carboxylic acid derivative
- Creates the first high-energy compound (acyl-phosphate bond in 1,3-BPG)
- Mechanism: GAPDH uses a cysteine residue to form a covalent thioester intermediate, conserving energy from the oxidation in the acyl-phosphate bond of 1,3-BPG. This is an example of covalent catalysis. - Basic Medical Biochemistry, p. 818
- This step requires NAD⁺. If NAD⁺ is not regenerated (e.g., no oxygen), glycolysis stops - this is why anaerobic regeneration of NAD⁺ (via lactate) is essential.
- Inhibited by iodoacetate (used in biochemistry research to block glycolysis)
2,3-Bisphosphoglycerate (2,3-BPG) shunt: In red blood cells, 1,3-BPG can be converted to 2,3-BPG (by bisphosphoglycerate mutase). 2,3-BPG allosterically decreases the affinity of hemoglobin for O₂, shifting the oxygen-dissociation curve to the right (Bohr-related effect). 2,3-BPG re-enters glycolysis as 3-phosphoglycerate. This shunt sacrifices ATP but serves a vital O₂-delivery function.
STEP 7: 1,3-Bisphosphoglycerate → 3-Phosphoglycerate (3-PG)
Enzyme: Phosphoglycerate kinase (PGK)
Reaction: 1,3-BPG + ADP → 3-Phosphoglycerate + ATP
Energy: Produces 2 ATP (1 per molecule × 2); reversible - this is substrate-level phosphorylation
Key points:
- First ATP-generating step of glycolysis
- "Substrate-level phosphorylation" means phosphate is transferred directly from a metabolic intermediate to ADP (unlike oxidative phosphorylation which uses the ETC)
- This step recoup the 2 ATP invested in steps 1 and 3
STEP 8: 3-Phosphoglycerate → 2-Phosphoglycerate (2-PG)
Enzyme: Phosphoglycerate mutase
Reaction: 3-PG ⇌ 2-PG (phosphate group moved from C3 to C2)
Energy: No ATP; reversible
Key points:
- A simple isomerization (intramolecular phosphate transfer)
- 2,3-BPG serves as a cofactor in this reaction - it acts as an intermediate in the phosphate transfer mechanism
- Prepares the substrate for the next highly exergonic step
STEP 9: 2-Phosphoglycerate → Phosphoenolpyruvate (PEP)
Enzyme: Enolase
Reaction: 2-PG ⇌ PEP + H₂O (dehydration)
Energy: No ATP; reversible
Key points:
- Removal of water creates PEP, which contains an extremely high-energy enol phosphate bond (ΔG°' of hydrolysis ≈ -61.9 kJ/mol)
- This stored energy is released in the final step
- Enolase requires Mg²⁺ and Zn²⁺
- Inhibited by fluoride (used in blood collection tubes for glucose testing - gray-top tubes contain NaF to inhibit enolase)
STEP 10: Phosphoenolpyruvate → Pyruvate
Enzyme: Pyruvate kinase (PK)
Reaction: PEP + ADP → Pyruvate + ATP
Energy: Produces 2 ATP (1 per molecule × 2); irreversible
Key points:
- Second and final ATP-generating step; substrate-level phosphorylation
- Allosteric activator: Fructose 1,6-bisphosphate (feedforward activation - when PFK-1 is active, it signals PK to also be active)
- Allosteric inhibitor: ATP, alanine (signals adequate energy and amino acid supply)
- Hormonal regulation (liver PK isoenzyme only):
- Insulin activates liver PK (dephosphorylation)
- Glucagon inhibits liver PK (phosphorylation via PKA cascade) - to suppress glycolysis and favor gluconeogenesis during fasting
- Pyruvate kinase deficiency: Autosomal recessive disorder. Most common enzymatic cause of hemolytic anemia. RBCs are entirely dependent on glycolysis for ATP; without adequate PK, ATP is depleted, the Na/K-ATPase fails, and cells lyse.
Summary of All 10 Steps
| Step | Substrate → Product | Enzyme | ATP/NADH | Reversible? |
|---|---|---|---|---|
| 1 | Glucose → G6P | Hexokinase/Glucokinase | -1 ATP | No |
| 2 | G6P → F6P | Phosphoglucose isomerase | - | Yes |
| 3 | F6P → F1,6-BP | PFK-1 (rate-limiting) | -1 ATP | No |
| 4 | F1,6-BP → G3P + DHAP | Aldolase | - | Yes |
| 5 | DHAP → G3P | Triose phosphate isomerase | - | Yes |
| 6 | G3P → 1,3-BPG | GAPDH | +2 NADH | Yes |
| 7 | 1,3-BPG → 3-PG | Phosphoglycerate kinase | +2 ATP | Yes |
| 8 | 3-PG → 2-PG | Phosphoglycerate mutase | - | Yes |
| 9 | 2-PG → PEP | Enolase | - | Yes |
| 10 | PEP → Pyruvate | Pyruvate kinase | +2 ATP | No |
Net yield: -2 ATP + 2 ATP + 2 ATP = +2 ATP net, +2 NADH
The 3 irreversible steps (1, 3, 10) are the regulatory points. The same three reactions are bypassed in gluconeogenesis by different enzymes.
Regulation of Glycolysis
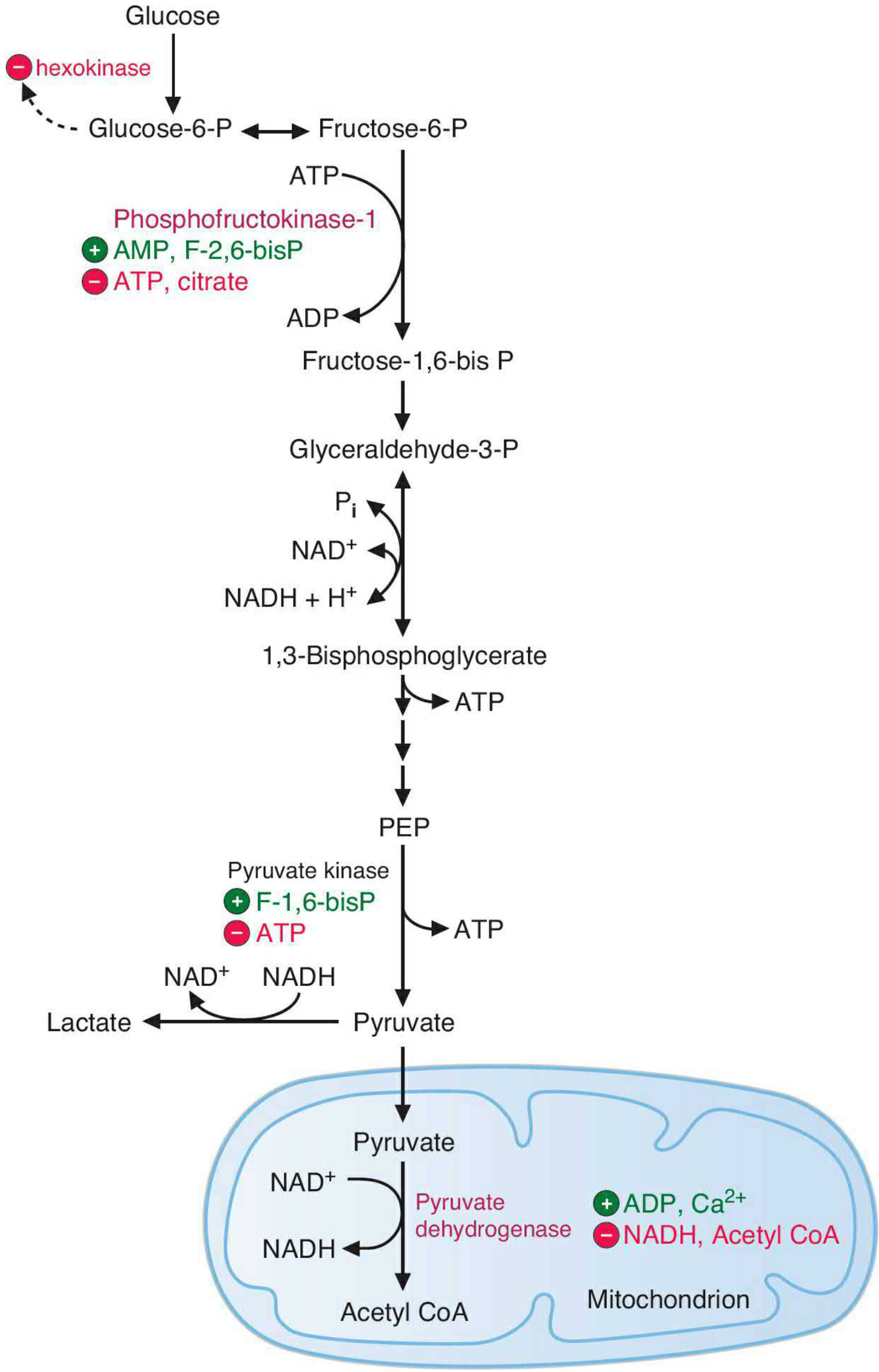
Figure: Major regulatory sites in glycolysis. Green circles = activators; red circles = inhibitors.
The pathway is primarily regulated to match ATP demand. - Basic Medical Biochemistry, p. 809
1. Hexokinase (Step 1)
- Inhibited by G6P (product inhibition) - prevents glucose sequestration when it is not needed
- Glucokinase: regulated by GKRP in response to glucose and fructose-6-P levels
2. PFK-1 (Step 3) - The Main Control Point
- Inhibited by: High ATP (signals energy sufficiency), citrate (TCA cycle backed up)
- Activated by: AMP, ADP (energy deficiency signal), F-2,6-BP (most potent activator), Pi
- The AMP:ATP ratio is the fundamental energy-sensing signal
- F-2,6-BP is the key hormonal messenger:
- Insulin → activates PFK-2 → increases F-2,6-BP → activates PFK-1 → glycolysis ON
- Glucagon → activates fructose 2,6-bisphosphatase → decreases F-2,6-BP → glycolysis OFF
3. Pyruvate Kinase (Step 10)
- Activated by: F-1,6-BP (feedforward activation from step 3)
- Inhibited by: ATP, alanine (signals energy surplus and amino acid availability)
- Liver isoenzyme additionally regulated by covalent phosphorylation: glucagon inhibits, insulin activates
AMP as a Sensitive Energy Signal
The adenylate kinase reaction (2 ADP ⇌ ATP + AMP) amplifies small changes in ATP. When ATP falls by 10%, AMP can rise dramatically, making AMP a far more sensitive indicator of falling energy charge than ATP itself. - Basic Medical Biochemistry, p. 811
Tissue-Specific Isoenzymes
All three regulatory enzymes exist as tissue-specific isoforms, tailoring glycolytic regulation to the function of each tissue. Liver isoforms, for example, are tuned to both glycolysis and gluconeogenesis needs.
Fate of Pyruvate
Pyruvate sits at a metabolic crossroads:
1. Aerobic conditions (mitochondria present, O₂ available)
- Pyruvate enters the mitochondria via the mitochondrial pyruvate carrier
- Converted to Acetyl-CoA by pyruvate dehydrogenase complex (PDC):
- Pyruvate + NAD⁺ + CoA → Acetyl-CoA + CO₂ + NADH
- Acetyl-CoA feeds the TCA (Krebs) cycle → oxidative phosphorylation → up to ~30-32 ATP total per glucose
2. Anaerobic conditions (no O₂, or no mitochondria)
- Pyruvate is reduced to lactate by lactate dehydrogenase (LDH):
- Pyruvate + NADH + H⁺ → Lactate + NAD⁺
- This regenerates NAD⁺, allowing glycolysis to continue
- No additional ATP is gained
- Net yield: only 2 ATP per glucose
3. In yeast and some microorganisms
- Pyruvate → Acetaldehyde + CO₂ (by pyruvate decarboxylase)
- Acetaldehyde → Ethanol + NAD⁺ (by alcohol dehydrogenase)
- This is fermentation
4. In liver (biosynthetic precursor)
- Pyruvate → Oxaloacetate (by pyruvate carboxylase) → gluconeogenesis
- Pyruvate → Acetyl-CoA → fatty acid synthesis
Anaerobic Glycolysis and the Cori Cycle
When tissues lack oxygen or mitochondrial capacity (e.g., exercising muscle, RBCs), lactate is produced and exported into the blood. The liver takes up this lactate and converts it back to glucose via gluconeogenesis. This glucose is then released into the blood for reuse. This cycling of lactate and glucose between muscle (or RBCs) and liver is called the Cori cycle. - Basic Medical Biochemistry, p. 805
Clinical relevance of lactate:
- Measuring blood lactate is used to detect O₂ debt (inadequate tissue oxygenation)
- Lactic acidosis occurs when circulatory collapse (MI, pulmonary embolism, hemorrhage, septic shock) deprives tissues of O₂, forcing them to rely on anaerobic glycolysis. Lactate accumulates, dropping blood pH. - Biochemistry, 8th ed, Lippincott, p. 318
Energy Yield - Full Accounting
| Condition | ATP from Glycolysis | Total ATP per Glucose |
|---|---|---|
| Anaerobic | Net 2 ATP | 2 ATP |
| Aerobic | Net 2 ATP (glycolysis) + 2 NADH (yields ~5 ATP via ETC) | ~30-32 ATP (including TCA + ETC) |
The 2 NADH produced in glycolysis must shuttle their electrons into the mitochondria (since NADH cannot cross the inner mitochondrial membrane directly). Depending on which shuttle is used:
- Malate-aspartate shuttle (heart, liver): each NADH yields ~2.5 ATP
- Glycerol-3-phosphate shuttle (muscle, brain): each NADH yields ~1.5 ATP
Special Topics
The Warburg Effect (Aerobic Glycolysis in Cancer)
Cancer cells preferentially use glycolysis even in the presence of adequate oxygen ("aerobic glycolysis"). Although glycolysis generates far less ATP per glucose than oxidative phosphorylation, it can produce ATP at a faster rate and generates biosynthetic precursors (nucleotides, amino acids, lipids) needed for rapid cell proliferation. HIF-1 (hypoxia-inducible factor-1) upregulates glycolytic enzymes in hypoxic tumors and in some normoxic cancer cells. - Basic Medical Biochemistry, p. 808
HIF-1 and Hypoxia
In hypoxic conditions (e.g., COPD), HIF-1 is activated in virtually all tissues. It increases transcription of glycolytic enzymes including PFK-1, enolase, phosphoglycerate kinase, and others, boosting anaerobic ATP production capacity.
2,3-Bisphosphoglycerate and RBCs
Red blood cells divert some 1,3-BPG into 2,3-BPG via the bisphosphoglycerate mutase shunt. 2,3-BPG stabilizes the deoxy form of hemoglobin, decreasing O₂ affinity and promoting O₂ release to tissues. At altitude, RBCs increase 2,3-BPG production to compensate for lower O₂ levels.
Dental Caries
Oral bacteria (Lactobacilli, Streptococcus mutans) derive virtually all their energy from anaerobic glycolysis, converting dietary sugars to lactic acid. Below pH 5.5, this acid dissolves tooth enamel (hydroxyapatite), causing dental caries. - Basic Medical Biochemistry, p. 805
Key Diseases/Clinical Correlations
| Condition | Enzyme/Target Affected | Consequence |
|---|---|---|
| Pyruvate kinase deficiency | Pyruvate kinase | Hemolytic anemia (RBCs ATP-depleted) |
| Galactosemia | Galactose-1-phosphate uridyltransferase | Galactose accumulation; toxic to liver, brain, lens |
| Lactic acidosis | Inadequate O₂ delivery | Impaired ETC, forced anaerobic glycolysis, H⁺ accumulation |
| Cancer (Warburg effect) | Multiple glycolytic enzymes upregulated | Aerobic glycolysis preferred for biosynthetic needs |
| Type 2 diabetes therapy (SGLT2 inhibitors) | SGLT2 (glucose transporter in kidney) | Increases renal glucose excretion, lowers blood glucose |
Summary of Key Concepts
- Glycolysis = 10 steps, cytosol, universal - operates with or without oxygen
- Phase 1 (steps 1-5) invests 2 ATP to activate glucose; produces 2 G3P
- Phase 2 (steps 6-10) yields 4 ATP and 2 NADH per glucose
- Net yield: 2 ATP + 2 NADH + 2 pyruvate per glucose
- Three irreversible reactions (hexokinase, PFK-1, pyruvate kinase) are the regulatory checkpoints
- PFK-1 is the primary rate-limiting enzyme; activated by AMP/ADP/F-2,6-BP; inhibited by ATP/citrate
- Fate of pyruvate: Acetyl-CoA (aerobic) or lactate (anaerobic)
- Anaerobic glycolysis regenerates NAD⁺ via lactate and sustains ATP production without O₂
- Warburg effect: cancer cells prefer glycolysis even in O₂ (for biosynthetic precursors + faster ATP rate)
- Clinical correlations include PK deficiency, lactic acidosis, and the metabolic basis of dental caries
Sources: Biochemistry, 8th ed, Lippincott Illustrated Reviews; Basic Medical Biochemistry - A Clinical Approach, 6th ed; Harper's Illustrated Biochemistry, 32nd ed; Medical Physiology (Boron & Boulpaep)
This is a shared conversation. Sign in to Orris to start your own chat.