Explain in detail about T Cell maturation, differentiation and activation. Mention briefly about further downstream activation of other immune cells. From Abbas and other immunology, rheumatology textbooks and journals
Reading File
Finding Sources
Finding Sources
Finding Sources
Searching PubMed
T cell maturation differentiation activation
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Loading Image
Reading File
Reading File
Excellent - I now have very rich content from Abbas (Cellular & Molecular Immunology), Janeway's Immunobiology, and Roitt's Essential Immunology. Let me compile the comprehensive answer now.
T Cell Maturation, Differentiation, and Activation
Sources: Abbas - Cellular and Molecular Immunology (10e), Janeway's Immunobiology (10e), Roitt's Essential Immunology, Fishman's Pulmonary Diseases, Yamada's Gastroenterology
1. Overview and Thymic Origin
T lymphocytes originate from multipotent progenitors that arise in the fetal liver and, postnatally, in the bone marrow. These precursors enter the thymus by crossing the endothelium of postcapillary venules at the corticomedullary junction. In mice, thymic seeding begins on embryonic day 11; in humans this corresponds to weeks 7-8 of gestation. The thymus is the primary site of T cell maturation - it involutes after puberty but a remnant persists, as evidenced by the successful immune reconstitution seen in adult bone marrow transplant recipients.
The thymic microenvironment is structurally organized to support sequential maturation events:
- Cortex: Thymic cortical epithelial cells form a meshwork of long cytoplasmic processes, expressing class I and II MHC molecules - the substrate for positive selection.
- Corticomedullary junction: Bone marrow-derived dendritic cells (DCs) reside here.
- Medulla: Medullary thymic epithelial cells (mTECs) and macrophages, critical for negative selection.
Migration through this compartmentalized anatomy is chemokine-driven: precursors use CCR9/CCL25 to enter the cortex; CCR7 (binding CCL21/CCL19) drives migration toward the medulla; mature thymocytes exit via the sphingosine-1-phosphate (S1P)/S1PR1 gradient into circulation. IL-7, secreted by thymic stromal cells, is the critical survival and proliferation cytokine throughout early maturation.
(Abbas - Cellular and Molecular Immunology, p. 603-604)
2. Stages of Thymocyte Maturation
2.1 Double-Negative (DN) Stage
The most immature thymocytes - recent bone marrow emigrants - express neither CD4 nor CD8. These double-negative (DN) thymocytes have TCR genes in germline configuration, with no surface TCR, CD3, or ζ chains. DN cells pass through four sub-stages (DN1-DN4) defined by CD44/CD25 expression:
- DN1/Pro-T cell: CD44+CD25−; not yet T-lineage committed; initiates migration to the mid-cortex.
- DN2/Pro-T: CD44+CD25+; committed to T lineage; RAG1 and RAG2 proteins are first expressed here, initiating TCR gene rearrangement. Dβ-to-Jβ rearrangements at the TCR β locus occur first.
- DN3/Pre-T: CD44−CD25+; Vβ-to-DJβ rearrangement completes. If a productive TCR β rearrangement occurs, the β chain pairs with the invariant pre-Tα chain to form the pre-TCR complex.
- DN4: The pre-TCR signals β-selection - confirming a productive rearrangement - and drives rapid proliferative expansion (a single precursor yields many progeny). It also suppresses further β chain rearrangements (allelic exclusion) and silences the γδ TCR program.
About 90% of DN cells productively rearrange TCR β first and are directed toward the αβ lineage. The remaining ~10% productively rearrange TCR γ and δ before TCR β and commit to the γδ lineage.
(Abbas, p. 605-607)
2.2 Double-Positive (DP) Stage
After β-selection, cells upregulate both CD4 and CD8 to become double-positive (DP) thymocytes, the largest population (~80%) in the thymus. At this stage:
- RAG1/RAG2 are re-expressed and TCR α chain rearrangement occurs (Vα-to-Jα joining).
- The complete αβ TCR complex assembles and is expressed on the surface with CD3 and ζ chains.
- Rearrangement of TCR α permanently deletes the TCR δ locus (embedded within TCR α), irreversibly committing the cell to the αβ lineage.
- DP thymocytes then undergo the critical selection processes.
(Abbas, p. 609-611)
3. Thymic Selection
3.1 Positive Selection
Positive selection occurs in the thymic cortex. DP thymocytes whose αβ TCRs bind with low/intermediate avidity to self peptide-MHC complexes on cortical epithelial cells receive a survival signal. This prevents death by neglect (default apoptosis for ~95% of thymocytes that fail to receive adequate signals).
The key outcomes of positive selection:
- MHC class I recognition → CD8+ single-positive T cell: TCR-class I interaction activates RUNX3, which maintains CD8 expression and silences CD4.
- MHC class II recognition → CD4+ single-positive T cell: Stronger signaling through CD4-LCK activates GATA3, which commits to the CD4 fate and induces ThPoK (a repressor of CD8 lineage genes).
The favored "instructional model" holds that the strength and quality of TCR signaling - influenced by coreceptor affinity for LCK (CD4 > CD8) - actively instructs lineage commitment rather than it being stochastic.
(Abbas, p. 612-613)
3.2 Negative Selection
Thymocytes whose TCRs bind self peptide-MHC complexes with high avidity undergo apoptosis via negative selection. This is the primary mechanism of central tolerance. It occurs mainly at the corticomedullary junction and in the thymic medulla, where mTECs and DCs present a diverse repertoire of self antigens.
A central molecular player is the AIRE (Autoimmune Regulator) transcription factor, expressed in mTECs. AIRE drives ectopic expression of peripheral tissue-specific antigens (insulin, thyroglobulin, etc.) in the medulla, enabling deletion of autoreactive T cells that would otherwise escape to the periphery. Loss-of-function AIRE mutations cause autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED), demonstrating the indispensability of this mechanism.
Cells that are neither positively nor negatively selected (the majority) die by neglect.
(Abbas, p. 614-618)
4. T Cell Activation: The Three-Signal Model
Naive T cells exiting the thymus recirculate through secondary lymphoid organs (lymph nodes, spleen, Peyer's patches), scanning APCs via the CCR7/CCL19-CCL21 axis. A naive T cell can survey approximately one lymphoid organ per day. Activation of a naive T cell - termed priming - requires at least three signals:
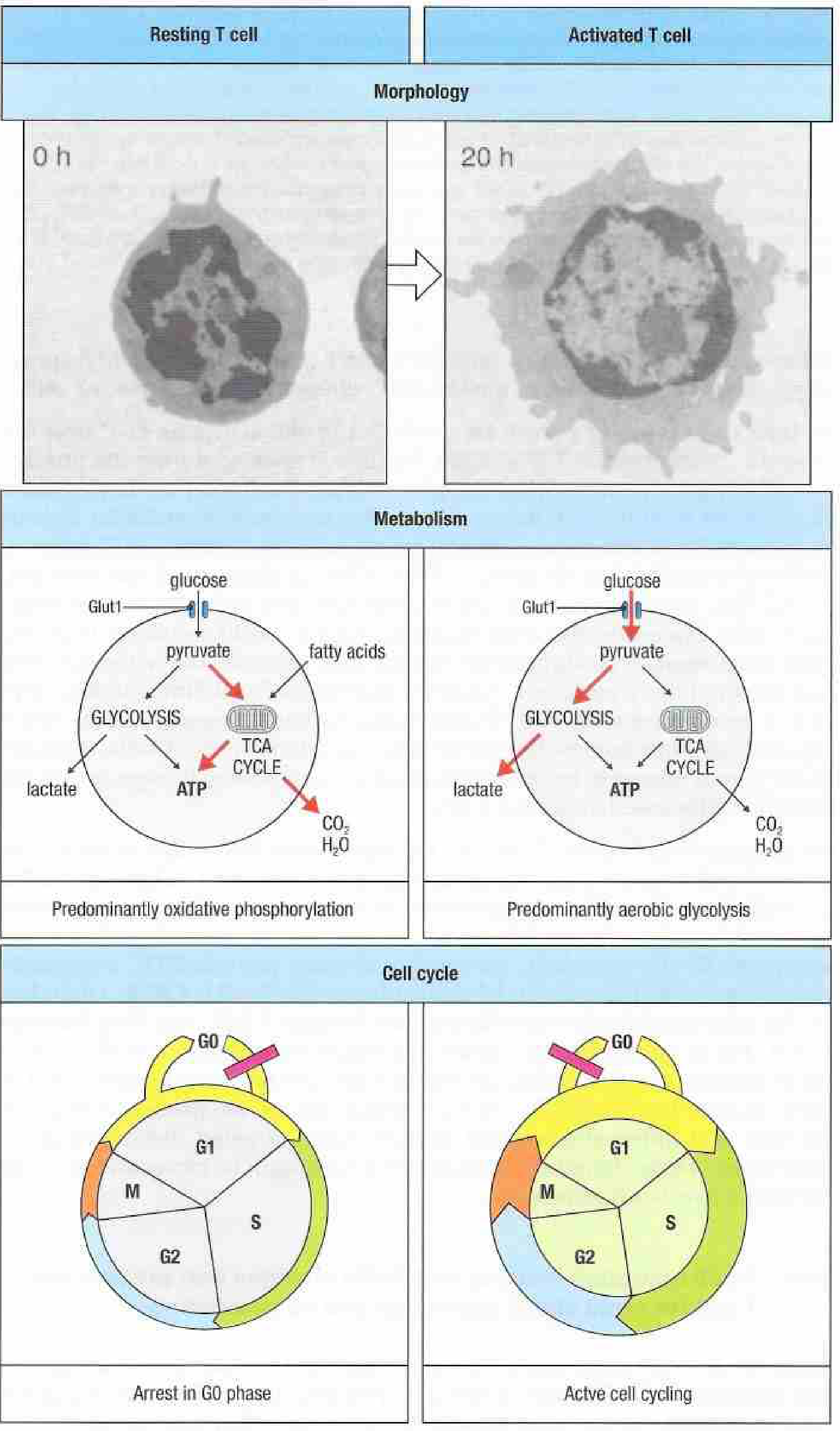
Signal 1 - TCR/Antigen Recognition
The TCR recognizes a specific peptide-MHC complex on an activated conventional dendritic cell (cDC). This is the antigen-specific primary signal. The CD4 or CD8 coreceptor binds the MHC molecule and brings LCK (a Src-family tyrosine kinase) into proximity with the TCR/CD3 complex.
Downstream TCR signaling cascade:
- LCK phosphorylates ITAMs (immunoreceptor tyrosine-based activation motifs) on CD3 ζ, γ, δ, ε chains.
- ZAP-70 (ζ-chain-associated protein kinase 70) is recruited and activated, phosphorylating the adaptor LAT (linker for activation of T cells).
- LAT recruits a signaling complex including PLCγ1 (phospholipase C gamma 1) and Grb2/SOS.
- PLCγ1 cleaves PIP2 into DAG (diacylglycerol) and IP3 (inositol triphosphate):
- IP3 → releases Ca²⁺ from ER → activates calcineurin → dephosphorylates and activates NFAT (Nuclear Factor of Activated T cells) → transcription of IL-2 and other effector genes.
- DAG → activates PKC-θ → activates IKK → releases NF-κB from IκB inhibition → NF-κB enters nucleus.
- Grb2/SOS activates Ras → MAPK/ERK cascade → activates AP-1 (Fos/Jun heterodimer) transcription factor.
The three transcription factors - NFAT, NF-κB, and AP-1 - act in a coordinated fashion at the IL-2 promoter and other cytokine gene promoters to initiate the gene expression program of T cell activation.
(Janeway's Immunobiology 10e, p. 387-390)
Signal 2 - Co-stimulation
TCR engagement alone (Signal 1 alone) induces T cell anergy - a state of unresponsiveness. Co-stimulation via CD28 (on T cells) binding B7-1 (CD80) or B7-2 (CD86) on APCs is required.
- CD28 signaling activates PI3K → Akt → mTOR, augmenting glucose uptake (Glut1 upregulation), promoting aerobic glycolysis (the Warburg-like metabolic shift), and driving cell cycle entry from G0 to G1.
- CD28 also stabilizes IL-2 mRNA and amplifies NFAT and NF-κB signals.
- Additional co-stimulatory pairs include ICOS/ICOS-L, OX40/OX40L, and 4-1BB/4-1BBL that reinforce or sustain activation.
- Negative co-stimulatory receptors CTLA-4 (CD152) and PD-1 (CD279) compete with or inhibit these signals to prevent uncontrolled activation.
(Janeway's, p. 389)
Signal 3 - Cytokine/Polarizing Signals
Cytokines produced by the APC and the innate immune environment determine the effector fate of the activated T cell. These signals act on STAT transcription factors and epigenetic regulators to lock in specific gene expression programs (see Section 5).
5. Effector T Cell Differentiation
Following priming, naive CD4+ T cells undergo clonal expansion (up to ~1000-fold for CD4; up to ~50,000-fold for CD8) over 2-3 days. This is fueled by the metabolic shift from oxidative phosphorylation to aerobic glycolysis (Warburg effect), which supplies biosynthetic precursors for nucleotides, lipids, and proteins needed for rapid cell division. The mTOR pathway (activated downstream of Akt) is the master regulator of this metabolic switch. Subsequently, the expanded clone undergoes clonal contraction (~90-95% die by apoptosis) after antigen clearance, leaving behind long-lived memory T cells.
5.1 CD4+ T Helper Cell Subsets
CD4+ naive T cells differentiate into distinct effector lineages defined by their cytokine milieu, master transcription factors, and effector cytokine outputs:
| Subset | Inducing Cytokines | Master TF | Signature Cytokines | Primary Function |
|---|---|---|---|---|
| Th1 | IL-12, IFN-γ | T-bet (TBX21) | IFN-γ, TNF-α, IL-2 | Macrophage activation; intracellular pathogens |
| Th2 | IL-4 | GATA3 (+ STAT6) | IL-4, IL-5, IL-13 | Eosinophil activation; helminth defense; allergy |
| Th17 | IL-6 + TGF-β (initiation); IL-23 (stabilization) | RORγt (+ STAT3) | IL-17A, IL-17F, IL-22 | Neutrophil recruitment; extracellular fungi/bacteria |
| Treg | TGF-β alone | FoxP3 | TGF-β, IL-10, IL-35 | Immune suppression; self-tolerance |
| Tfh | IL-6, IL-21 | Bcl-6 (+ CXCR5) | IL-21, IL-4 | Germinal center formation; B cell help |
| Th9 | IL-4 + TGF-β | PU.1, IRF4 | IL-9 | Mast cell activation; anti-tumor |
Th1 Differentiation:
IL-12 (from DCs and macrophages) signals via STAT4 to induce T-bet. IFN-γ (from NK cells or existing Th1 cells) signals via STAT1 and further reinforces T-bet expression. IL-12 and IFN-γ both downregulate IL-4 receptor signaling, creating a positive feedback loop for Th1 commitment.
Th2 Differentiation:
IL-4 (from NKT cells, basophils, and possibly the T cell itself) activates STAT6 → GATA3. GATA3 drives expression of IL-4, IL-5, and IL-13 while also downregulating the IL-12Rβ2 subunit - making the cell unresponsive to Th1-polarizing IL-12. IL-4 effects are dominant: the ratio of IL-4 to IL-12/IFN-γ in the environment is the deciding factor.
(Roitt's Essential Immunology, p. 268)
Th17 Differentiation:
IL-6 + TGF-β (in combination) activate STAT3 → RORγt - the master Th17 regulator. TGF-β alone drives Treg differentiation; IL-6 redirects TGF-β signaling toward Th17 by suppressing FoxP3 and inducing RORγt. IL-23 (acting via IL-23R, which is upregulated upon activation) then stabilizes and expands committed Th17 cells. Th17 cells produce IL-17A/F, IL-22, and IL-23, driving neutrophilic inflammation and mucosal defense. Pathologically, Th17 cells drive rheumatoid arthritis, psoriasis, ankylosing spondylitis, and inflammatory bowel disease.
(Roitt's, p. 268-269)
Treg Differentiation:
TGF-β alone (without IL-6) activates FoxP3, the lineage-defining transcription factor of natural and induced Tregs. FoxP3+ Tregs suppress immune responses through: (a) secretion of TGF-β and IL-10; (b) CTLA-4-mediated competition for CD28 co-stimulation; (c) consumption of IL-2 (high CD25 expression acts as an IL-2 "sink"); (d) direct cytolysis via granzymes.
Tfh Differentiation:
Bcl-6 upregulation (driven by IL-6 and IL-21) is the central event. Bcl-6 upregulates CXCR5 (permitting follicular homing via CXCL13) and simultaneously represses T-bet, GATA3, RORγt, and FoxP3 - preventing commitment to all other lineages. Tfh cells provide B cell help for germinal center reactions.
(Roitt's, p. 269)
5.2 CD8+ Cytotoxic T Lymphocytes (CTLs)
Naive CD8+ T cells are preferentially activated by cDC1 dendritic cells (cross-presenters). Primed by Signal 1 (TCR/peptide-MHC I), Signal 2 (CD28), and Signal 3 (IL-12, type I IFNs), they differentiate into CTLs. CTLs kill target cells via:
- Perforin/granzyme pathway: Polarized exocytosis of cytotoxic granules; perforin creates pores in the target cell membrane, and granzymes B (key in humans) and A enter and activate caspases → apoptosis.
- Fas/FasL (CD95/CD95L) pathway: CTL surface FasL engages Fas on the target cell → DISC formation → caspase-8 activation → apoptosis.
- Cytokine secretion: IFN-γ and TNF-α produced by CTLs directly kill infected cells and activate macrophages.
6. Memory T Cells
Following clonal contraction, ~5-10% of the expanded clone survives as memory T cells. These are maintained by homeostatic cytokines (IL-7, IL-15) and persist for decades. Key memory populations:
- Central Memory (Tcm): CCR7+, CD62L+; reside in lymph nodes; rapidly proliferate on reencounter with antigen.
- Effector Memory (Tem): CCR7−, CD62L−; reside in peripheral tissues and blood; immediate effector function.
- Tissue-Resident Memory (Trm): CD69+, CD103+; permanently reside in tissues (lung, gut, skin); first-line responders.
- Memory Stem Cells (Tscm): CD45RA+, CCR7+; long-lived, highly self-renewing; the apex of memory hierarchy.
7. Downstream Activation of Other Immune Cells by T Cells
7.1 B Cell Activation and Antibody Production
Tfh cells migrate into B cell follicles (CXCR5/CXCL13), where they interact with antigen-primed B cells in the germinal center. This interaction involves:
- CD40L (CD154) on Tfh binding CD40 on B cells - the essential contact signal for B cell class switching and survival.
- Cytokines: IL-21 drives B cell proliferation and differentiation into plasma cells; IL-4 induces IgE class switching; IFN-γ drives IgG subclass switching.
- The GC reaction produces high-affinity antibody-secreting plasma cells and long-lived memory B cells.
Th1 cells also provide CD40L/IL-2 signals to B cells, driving predominantly IgG1/IgG3 class switching in humans.
7.2 Macrophage Activation
Th1-derived IFN-γ is the key classical macrophage activator:
- Upregulates MHC II, CD80/86, and iNOS.
- Induces the "classically activated" M1 macrophage phenotype, producing TNF-α, IL-12, IL-1β, and reactive oxygen/nitrogen species.
- Enhances intracellular killing (critical against Mycobacterium tuberculosis, Leishmania).
CD40L (from Th1 cells) binding CD40 on macrophages also provides a contact-dependent activating signal, synergizing with IFN-γ.
(Abbas, p. 611)
7.3 Neutrophil Recruitment
Th17-derived IL-17A/F and IL-22 act on stromal cells, epithelium, and endothelium to:
- Induce G-CSF and CXCL8 (IL-8) production → neutrophil mobilization and recruitment.
- Drive production of antimicrobial peptides (defensins, calprotectin) at mucosal barriers.
7.4 Eosinophil Activation and IgE-Mediated Immunity
Th2-derived IL-5 is the key eosinophil survival, differentiation, and activation cytokine. IL-4 and IL-13 act on B cells to switch to IgE production, which arms mast cells and basophils via FcεRI receptors.
7.5 NK Cell and Innate Cell Cross-talk
- IL-2 secreted by activated CD4+ T cells drives NK cell proliferation and enhances cytotoxicity.
- IFN-γ from T cells primes NK cells and macrophages.
- Conversely, NK cell-derived IFN-γ feeds back to reinforce Th1 differentiation.
7.6 Dendritic Cell Licensing
CD4+ T cells with CD40L can "license" DCs - converting an immature DC into a fully co-stimulatory APC capable of priming naive CD8+ T cells even without direct CD4 help at the time of CD8 priming (the "DC licensing" model of cross-priming). This is critical for antitumor immunity and viral infections.
8. Epigenetics and Metabolic Checkpoints
Roitt's emphasizes that T cell activation and lineage commitment are regulated at the epigenetic level:
- Pioneer transcription factors displace nucleosomes at gene enhancers (creating open chromatin before the transcription machinery arrives).
- Histone modifications: H3K4me3 (active) marks Th1-associated IFN-γ locus in Th1 cells; H3K27me3 methylation represses the same locus in Th2 cells.
- DNA methylation at CpG islands silences lineage-inappropriate cytokine genes.
- mTORC1 (activated by Akt downstream of CD28) drives aerobic glycolysis and is required for Th1 and Th17 differentiation; mTORC2 promotes Th2. Treg development preferentially uses fatty acid oxidation and OXPHOS rather than aerobic glycolysis - explaining why glucose deprivation (as in tumors) enriches for Tregs.
- HIF-1α, induced in low-oxygen environments, reinforces Th17 fate via direct activation of RORγt, while simultaneously targeting FoxP3 for proteasomal degradation - tilting the Th17/Treg balance toward inflammation.
(Roitt's, p. 220-222, 268)
Summary Table: Key T Cell Maturation Checkpoints
| Stage | Location | Key Event | Selection Type |
|---|---|---|---|
| DN (pre-TCR) | Cortex | β-chain rearrangement; pre-TCR checkpoint | β-selection |
| DP | Cortex | αβ TCR expression; cortical epithelium interaction | Positive selection |
| DP → SP transition | Cortex/medulla junction | Lineage commitment (CD4 vs CD8) | - |
| SP in medulla | Medulla | High-avidity self-antigen recognition | Negative selection (AIRE) |
| Naive T cell | Periphery | APC encounter; 3-signal activation | Antigen-driven |
| Effector differentiation | Lymph node | Cytokine-driven lineage commitment (Th1/2/17/Treg/Tfh/CTL) | - |
| Memory formation | Periphery/tissues | Homeostatic survival (IL-7, IL-15) | - |
Key references:
- Abbas AK, Lichtman AH, Pillai S. Cellular and Molecular Immunology, 10th ed. - Chapters 8 (T cell development), 9 (T cell activation), 10 (effector T cells)
- Murphy K, Weaver C. Janeway's Immunobiology, 10th ed. - Chapters 8, 9 (T cell signaling and activation)
- Roitt I, Delves P, Martin S, Burton D. Roitt's Essential Immunology, 13th ed. - Chapters 7-8 (T cell activation, metabolic reprogramming, Th polarization)
- Yamada T et al. Yamada's Textbook of Gastroenterology, 7th ed. - T cell lineages in mucosal immunity
- Fishman AP et al. Fishman's Pulmonary Diseases and Disorders, 5th ed. - Adaptive T cell responses in lung immunity
This is a shared conversation. Sign in to Orris to start your own chat.