Substrat level phosphorylation
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Here is a thorough breakdown of substrate-level phosphorylation (SLP):
Substrate-Level Phosphorylation (SLP)
Definition
Substrate-level phosphorylation is the formation of a high-energy phosphate bond where none previously existed, without the use of molecular O₂ - that is, it does not require the electron transport chain (ETC) or oxidative phosphorylation. Instead, phosphate is transferred directly from a high-energy phosphorylated intermediate (the "substrate") to ADP, producing ATP (or GTP).
"The transfer of phosphate from high-energy intermediates of the pathway to ADP" - Basic Medical Biochemistry, A Clinical Approach
It contrasts with oxidative phosphorylation, which uses the proton gradient generated by the ETC and requires oxygen.
Where It Occurs
1. Glycolysis (Cytoplasm) - 4 ATP produced, net 2 ATP
Two reactions in glycolysis generate ATP by SLP:
| Reaction | Enzyme | High-Energy Substrate | ATP Produced per Glucose |
|---|---|---|---|
| 1,3-Bisphosphoglycerate → 3-Phosphoglycerate | Phosphoglycerate kinase | Acyl phosphate bond (~10 kcal/mol) | 2 ATP |
| Phosphoenolpyruvate (PEP) → Pyruvate | Pyruvate kinase | Enolphosphate bond (~14 kcal/mol) | 2 ATP |
Key mechanism: Glyceraldehyde 3-phosphate dehydrogenase first oxidizes glyceraldehyde 3-P to form the high-energy acyl phosphate in 1,3-bisphosphoglycerate. This "primes the pump" - the oxidation energy is stored as a high-energy phosphate bond that can then be transferred to ADP. This is the entry point into SLP in glycolysis.
Net ATP from SLP in glycolysis = 2 ATP per glucose (4 produced minus 2 consumed in the investment phase).
2. TCA Cycle (Mitochondrial Matrix) - 1 GTP per turn
| Reaction | Enzyme | Product |
|---|---|---|
| Succinyl-CoA → Succinate | Succinate thiokinase (succinyl-CoA synthetase) | 1 GTP (equivalent to 1 ATP) |
Mechanism: The energy of the high-energy thioester bond of succinyl-CoA is used to phosphorylate GDP to GTP. The GTP produced is energetically equivalent to ATP and can be used directly in protein synthesis and other energy-requiring processes.
Per turn of the TCA cycle = 1 GTP from SLP.
Mechanism - How Is a High-Energy Bond Created?
The key insight is that SLP requires a prior oxidation step to "create" energy in the first place:
- An oxidation reaction releases energy
- That energy is captured as a high-energy phosphate bond (rather than lost as heat)
- The high-energy bond is then transferred to ADP
This is why glyceraldehyde 3-P dehydrogenase (an oxidative enzyme) is so central to glycolytic SLP - the oxidation energy is funneled directly into bond formation.
Comparison: SLP vs Oxidative Phosphorylation
| Feature | Substrate-Level Phosphorylation | Oxidative Phosphorylation |
|---|---|---|
| Location | Cytoplasm (glycolysis), mitochondrial matrix (TCA) | Inner mitochondrial membrane |
| Oxygen required? | No | Yes |
| Mechanism | Direct phosphate transfer from substrate to ADP | Proton gradient drives ATP synthase |
| ATP yield | Low (2-3 ATP/glucose) | High (~28-30 ATP/glucose) |
| Key enzymes | Phosphoglycerate kinase, Pyruvate kinase, Succinate thiokinase | ATP synthase (Complex V) |
| Occurs in anaerobic conditions? | Yes | No |
| Product | ATP (or GTP) | ATP |
Clinical Relevance
-
Anaerobic conditions (e.g., intense exercise, ischemia): When oxygen is unavailable, oxidative phosphorylation ceases. SLP in glycolysis becomes the only source of ATP, though its yield is limited (only 2 net ATP/glucose vs ~30-32 from full aerobic oxidation).
-
Cancer cells (Warburg effect): Many cancer cells rely heavily on glycolysis even in the presence of oxygen ("aerobic glycolysis"), depending on SLP for rapid (though inefficient) ATP generation to support rapid proliferation.
-
Cyanide/CO poisoning: These block the ETC, completely stopping oxidative phosphorylation. The cell falls back entirely on SLP, which cannot maintain adequate energy supply - leading to cellular energy failure.
-
Red blood cells (RBCs): Have no mitochondria and depend exclusively on glycolytic SLP for all their ATP needs.
ATP Count Summary
| Pathway | Reactions with SLP | ATP/GTP per Glucose |
|---|---|---|
| Glycolysis | Phosphoglycerate kinase + Pyruvate kinase | 2 ATP net |
| TCA cycle (×2 turns per glucose) | Succinate thiokinase | 2 GTP |
| Total from SLP | ~4 ATP equivalents |
The remaining ~28 ATP come from oxidative phosphorylation via NADH and FADH₂ generated in glycolysis and the TCA cycle.
Sources: Basic Medical Biochemistry - A Clinical Approach, 6e; Biochemistry, 8th ed - Lippincott Illustrated Reviews; Harper's Illustrated Biochemistry, 32nd Ed
Tca cycle
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Loading Image
Loading Image
Here is a complete guide to the TCA cycle from your biochemistry textbooks:
The TCA Cycle (Tricarboxylic Acid / Krebs / Citric Acid Cycle)
Overview
The TCA cycle is a series of 8 reactions in the mitochondrial matrix that oxidizes the acetyl group (2C) of acetyl-CoA to CO₂, capturing energy as NADH, FADH₂, and GTP. It is the final common pathway for oxidation of carbohydrates, fats, and proteins. It is an aerobic pathway - it requires oxygen indirectly because it depends on NAD⁺/FAD being continuously regenerated by the respiratory chain.
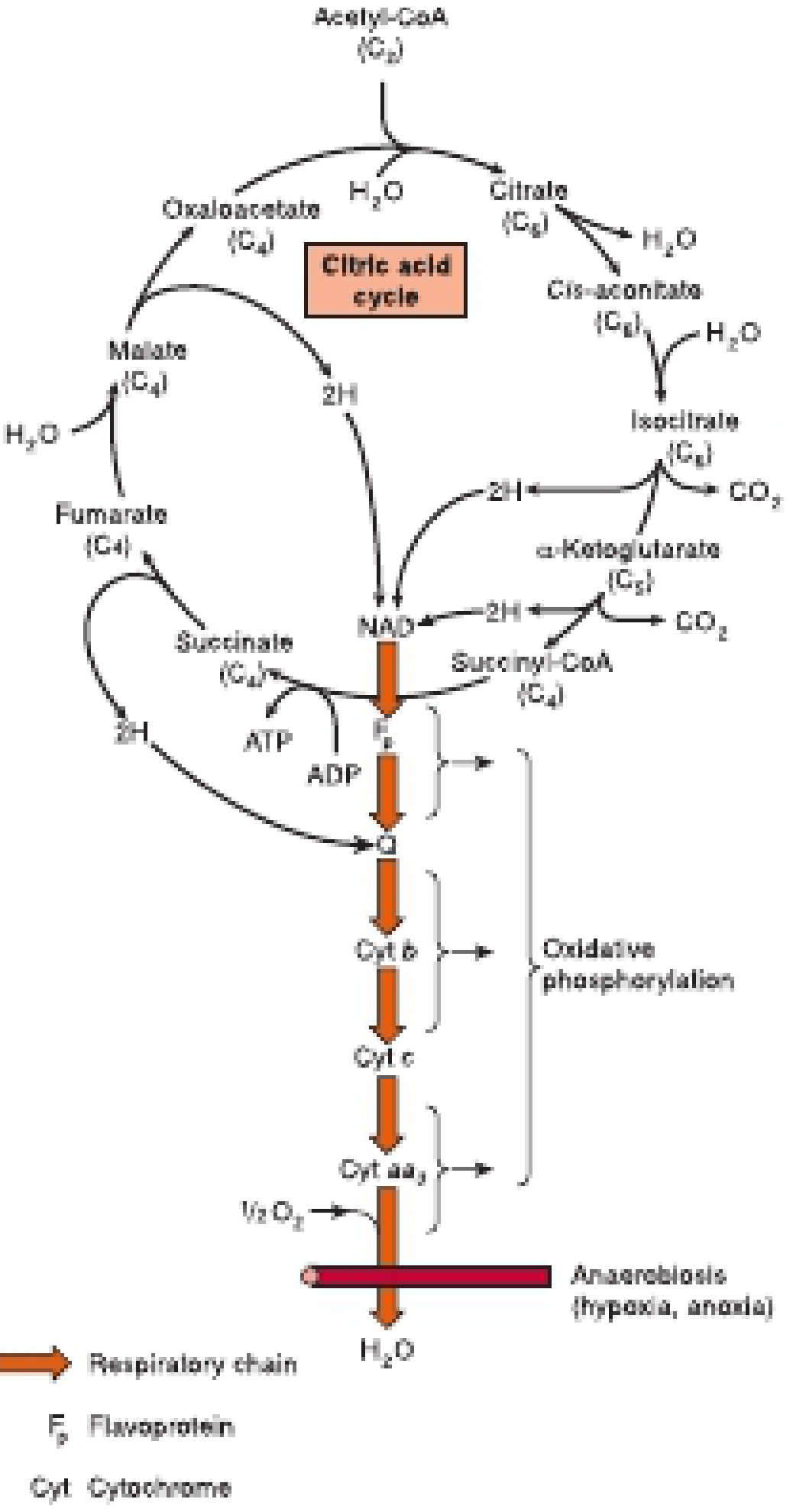
Entry Point - Pyruvate Dehydrogenase Complex
Before entering the TCA cycle, pyruvate (from glycolysis) is converted to acetyl-CoA in the mitochondrial matrix:
Pyruvate + CoA + NAD⁺ → Acetyl-CoA + CO₂ + NADH
Cofactors required: Thiamin (B1), Lipoate, CoA, FAD (B2), NAD⁺ (B3) - remembered as "TLC FAN"
The 8 Reactions
Here is the detailed reaction diagram from Harper's Biochemistry:
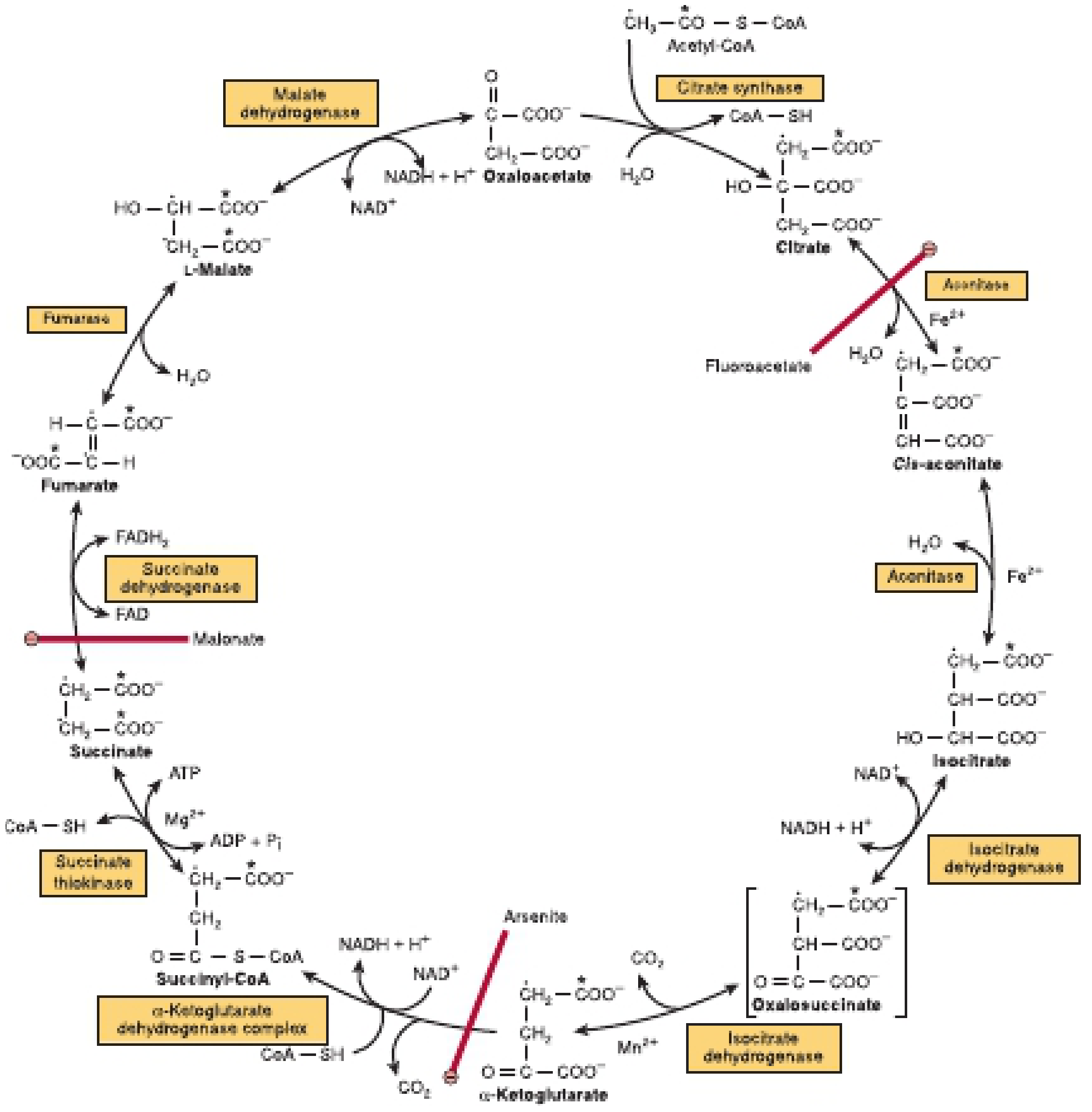
| Step | Substrate → Product | Enzyme | Coenzyme/Product | Notes |
|---|---|---|---|---|
| 1 | Oxaloacetate + Acetyl-CoA → Citrate | Citrate synthase | CoA released | Irreversible; rate-limiting step; inhibited by ATP, NADH, citrate |
| 2 | Citrate → Isocitrate | Aconitase | Requires Fe²⁺; intermediate cis-aconitate | Inhibited by fluoroacetate (via fluorocitrate) |
| 3 | Isocitrate → α-Ketoglutarate + CO₂ | Isocitrate dehydrogenase | NADH produced; requires Mn²⁺/Mg²⁺ | First CO₂ released; activated by ADP, Ca²⁺; inhibited by ATP, NADH |
| 4 | α-Ketoglutarate → Succinyl-CoA + CO₂ | α-Ketoglutarate dehydrogenase complex | NADH produced | Second CO₂ released; inhibited by arsenite, succinyl-CoA, NADH; same cofactors as PDH |
| 5 | Succinyl-CoA → Succinate | Succinate thiokinase (succinyl-CoA synthetase) | GTP (substrate-level phosphorylation) | Only SLP step in TCA; uses energy of thioester bond |
| 6 | Succinate → Fumarate | Succinate dehydrogenase | FADH₂ produced | Embedded in inner mitochondrial membrane; inhibited by malonate (competitive); directly reduces ubiquinone (Q) |
| 7 | Fumarate → L-Malate | Fumarase (fumarate hydratase) | H₂O added | Stereospecific - only produces L-malate |
| 8 | L-Malate → Oxaloacetate | Malate dehydrogenase | NADH produced | Equilibrium favors malate, but OAA is rapidly consumed, driving reaction forward |
Products Per Turn (per Acetyl-CoA)
| Product | Quantity | ATP equivalents |
|---|---|---|
| NADH | 3 | 3 × 2.5 = 7.5 ATP |
| FADH₂ | 1 | 1 × 1.5 = 1.5 ATP |
| GTP | 1 | 1 ATP |
| CO₂ | 2 | - |
| Total | ~10 ATP per turn |
Per glucose molecule, the TCA cycle runs twice (2 acetyl-CoA produced) = ~20 ATP from TCA alone.
Regulation
The cycle is primarily regulated by availability of NAD⁺ (which depends on ADP availability and ATP demand - respiratory control). Key regulatory enzymes:
| Enzyme | Activators | Inhibitors |
|---|---|---|
| Citrate synthase | Oxaloacetate | ATP, NADH, citrate, long-chain fatty acyl-CoA, succinyl-CoA |
| Isocitrate dehydrogenase | ADP, Ca²⁺ | ATP, NADH |
| α-Ketoglutarate dehydrogenase | Ca²⁺ | Succinyl-CoA, NADH, ATP; arsenite (blocks lipoate); NH₃ (in liver disease) |
Ca²⁺ is a key activator during muscle contraction - it simultaneously activates multiple dehydrogenases to match energy demand.
Vitamins Required by the TCA Cycle
| Vitamin | Coenzyme form | Role in TCA |
|---|---|---|
| B1 (Thiamin) | Thiamin diphosphate | α-Ketoglutarate dehydrogenase, PDH |
| B2 (Riboflavin) | FAD | Succinate dehydrogenase |
| B3 (Niacin) | NAD⁺ | Isocitrate DH, α-KG DH, Malate DH |
| Pantothenic acid (B5) | Coenzyme A | Acetyl-CoA, Succinyl-CoA formation |
Amphibolic Role - Beyond Energy Production
The TCA cycle is amphibolic - it serves both catabolic AND anabolic roles:
Anaplerosis (refilling the cycle):
- Pyruvate → Oxaloacetate (pyruvate carboxylase; requires biotin/B7)
- Amino acids (glutamate → α-ketoglutarate; aspartate → oxaloacetate; etc.)
Cataplerosis (draining the cycle for biosynthesis):
- Citrate → cytosol → acetyl-CoA (for fatty acid synthesis)
- Oxaloacetate → PEP (gluconeogenesis)
- α-Ketoglutarate → glutamate → amino acids
- Succinyl-CoA → heme synthesis
Clinical Connections
| Condition | Mechanism | Effect on TCA |
|---|---|---|
| Thiamin (B1) deficiency (Wernicke's, beri-beri) | α-KG DH and PDH impaired | TCA flux reduced; α-ketoglutarate accumulates; lactic acidosis |
| Arsenite poisoning | Inhibits lipoate-dependent enzymes (α-KG DH, PDH) | Blocks TCA at step 4; α-KG accumulates |
| Fluoroacetate poisoning | Forms fluorocitrate → inhibits aconitase | TCA blocked at step 2; citrate accumulates |
| Malonate poisoning | Competitive inhibitor of succinate dehydrogenase | TCA blocked at step 6; succinate accumulates |
| Liver disease / Hyperammonemia | NH₃ inhibits α-KG DH; withdrawal of α-KG to form glutamine | TCA depleted; ATP falls; coma |
| Hypoxia/Ischemia | NAD⁺ not regenerated (no ETC) | TCA stops; cell energy failure |
Sources: Harper's Illustrated Biochemistry, 32nd Ed; Guyton & Hall Textbook of Medical Physiology; Basic Medical Biochemistry - A Clinical Approach, 6e
This is a shared conversation. Sign in to Orris to start your own chat.