Biochemistry of inner ear fluids and potassium transport. MS ENT university pattern answer based on standard ENT textbooks.
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
I now have all the content and diagrams needed. Let me compose the comprehensive MS ENT university pattern answer.
Biochemistry of Inner Ear Fluids and Potassium Transport
Sources: Cummings Otolaryngology Head and Neck Surgery (7th ed); Shambaugh's Surgery of the Ear (6th ed); Bradley & Daroff's Neurology in Clinical Practice
I. Introduction
The inner ear contains two distinct fluid compartments - perilymph and endolymph - that differ dramatically in their ionic composition. This unique biochemical arrangement generates the endocochlear potential (ECP), an electrical driving force of approximately +85 mV that is indispensable for mechanosensory transduction in hair cells. Understanding these fluids and the potassium transport machinery that sustains them is fundamental to understanding auditory physiology, hereditary deafness, and ototoxicity.
II. Structural Compartments of the Cochlea
The cochlea is divided into three fluid-filled compartments by membranous partitions:
| Compartment | Bounding Structures | Fluid |
|---|---|---|
| Scala vestibuli | Above Reissner's membrane | Perilymph |
| Scala media (cochlear duct) | Between Reissner's membrane and basilar membrane | Endolymph |
| Scala tympani | Below basilar membrane | Perilymph |
The scala vestibuli and scala tympani connect at the apex via the helicotrema. The scala media contains the organ of Corti resting on the basilar membrane, and the stria vascularis lines its lateral wall.
III. Biochemical Composition of Inner Ear Fluids
A. Perilymph
Perilymph fills the scala vestibuli and scala tympani. Its ionic composition closely resembles extracellular fluid:
| Ion | Concentration |
|---|---|
| Na+ | ~140 mEq/L (high) |
| K+ | ~5-7 mEq/L (low) |
| Ca2+ | ~1.2 mM (normal ECF levels) |
| Protein | Low (~0.3 g/dL) |
Origin: Perilymph is derived primarily from blood plasma via ultrafiltration, with a minor contribution from CSF via the cochlear aqueduct. Its high Na+/low K+ profile reflects its extracellular character.
B. Endolymph
Endolymph fills the scala media (and the membranous labyrinth of the vestibular organs). Its ionic composition is unique among extracellular fluids - it closely resembles intracellular fluid:
| Ion | Concentration |
|---|---|
| K+ | ~150 mEq/L (very high) |
| Na+ | ~1-2 mEq/L (very low) |
| Ca2+ | ~20 µM (unusually low) |
| Protein | Very low |
This high-K+/low-Na+/low-Ca2+ signature is extraordinary - no other extracellular fluid in the body has this composition. Its maintenance depends entirely on active cellular transport mechanisms, chiefly in the stria vascularis.
Origin: Endolymph is actively secreted by the stria vascularis; it is not a passive ultrafiltrate. The endolymphatic sac participates in its reabsorption and volume regulation.
C. Intrastrial Fluid
A third, less-discussed compartment exists within the stria vascularis itself - the intrastrial compartment - sandwiched between the marginal cell layer and the basal cell layer:
- High Na+, low K+ (similar to perilymph)
- Electrical potential of approximately +90 to +100 mV
- Isolated from both endolymph and perilymph by tight junctions
IV. The Endocochlear Potential (ECP)
The scala media maintains a resting electrical potential of +60 to +100 mV (classically cited as +85 mV) relative to perilymph. This is the ECP (also called the endolymphatic potential).
Significance of the ECP:
- It adds to the hair cell's resting intracellular potential of approximately -45 mV
- The total electrochemical driving force across the apical hair cell membrane is therefore ~130 mV (+85 mV ECP + 45 mV intracellular negativity)
- This large driving force enables rapid influx of K+ (and Ca2+) through mechanotransduction channels with minimal metabolic cost to the hair cell itself
- Hearing threshold increases approximately 1 dB per mV loss of ECP (Shambaugh)
The ECP is generated and maintained exclusively by the stria vascularis. It is abolished by furosemide (loop diuretic), which inhibits NKCC1, and by ouabain, which inhibits Na+/K+-ATPase.
V. The Stria Vascularis - Structure and Function
The stria vascularis is a highly vascularized, multilayered tissue on the outer wall of the scala media. It is the "battery" of the cochlea, responsible for maintaining endolymph composition and the ECP.
Three cell layers (from endolymph side to perilymph side):
- Marginal cells - face the scala media (endolymph); actively secrete K+ into endolymph
- Intermediate cells - embedded in the intrastrial compartment alongside blood vessels; release K+ into intrastrial space via KCNJ10 channels
- Basal cells - face the spiral ligament/perilymph; connected by gap junctions to fibrocytes of spiral ligament
Tight junctions at the marginal cell layer and the basal cell layer create ionic barriers that define the intrastrial compartment. The intermediate and basal cells are electrically coupled via gap junctions.
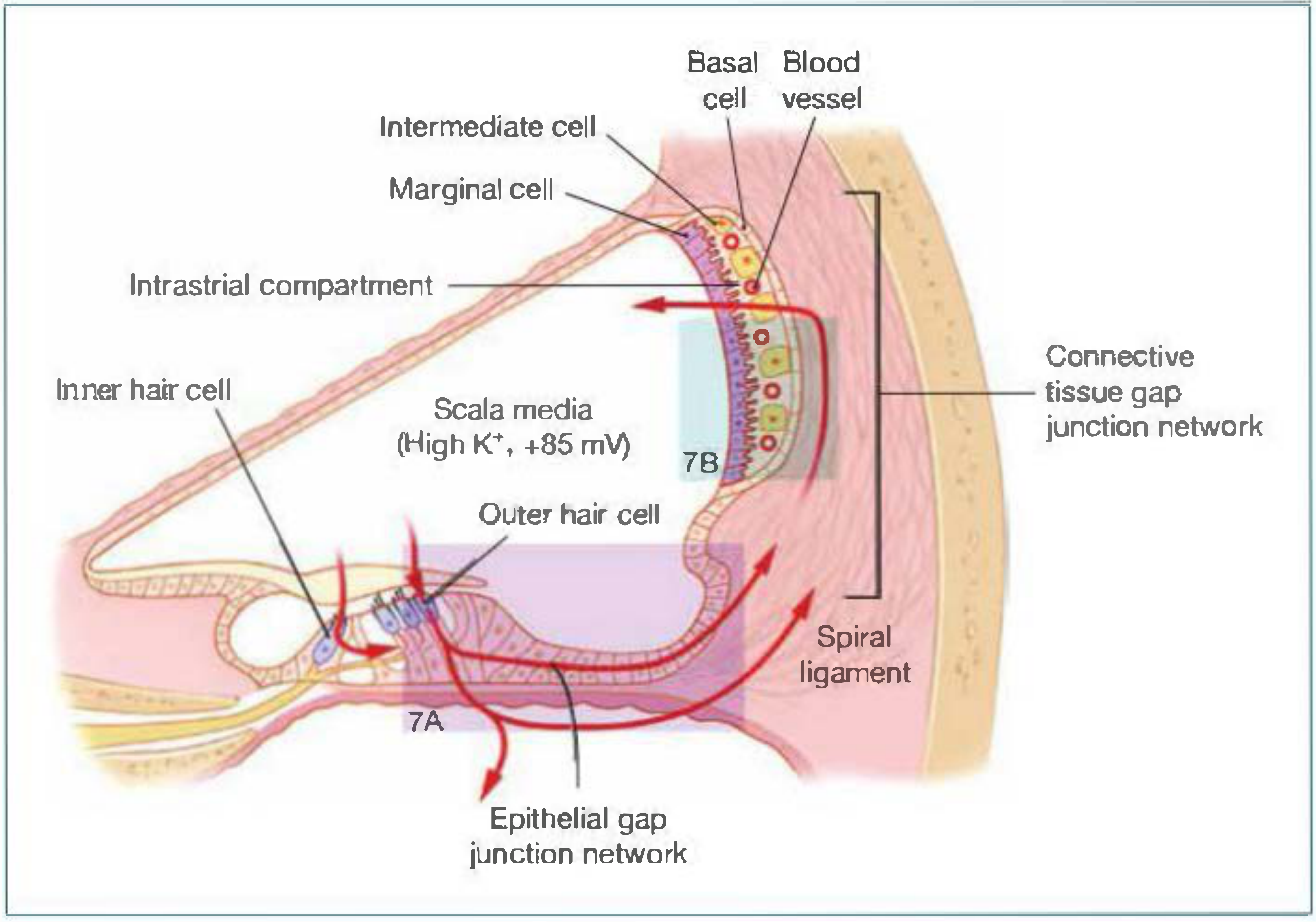
Fig. 1 - K+ circulation through the cochlea (Shambaugh's Surgery of the Ear, Fig. 4-6)
VI. Potassium Transport - The K+ Recycling Circuit
K+ is the principal cation mediating mechanotransduction. After entering hair cells from the endolymph, K+ must be returned to the stria vascularis to replenish endolymph. This "K+ recycling circuit" (or more accurately, K+ circulation pathway) operates through two parallel routes:
The Complete K+ Circuit (Step-by-Step)
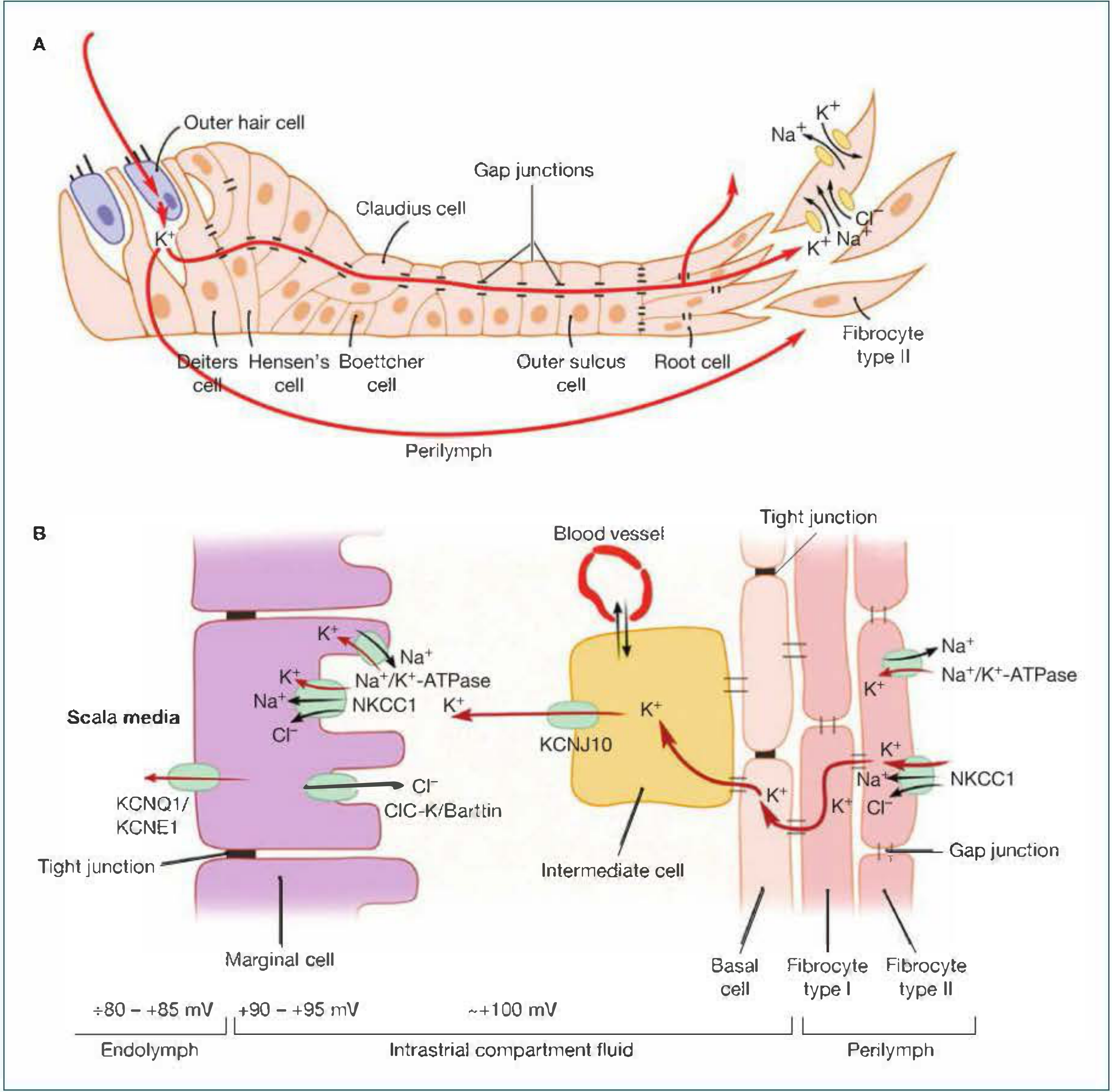
Fig. 2 - Detailed K+ transport at cellular level (Shambaugh's Surgery of the Ear, Fig. 4-7)
Step 1 - Entry into hair cells (apical surface):
- K+ (and Ca2+) enters hair cells through mechanotransduction channels (TMC1/TMC2) at the tips of stereocilia
- Driven by the large electrochemical gradient (+130 mV total driving force)
- Deflection toward tallest stereocilia = tip link stretch = channel opening = K+ influx = depolarization
- Deflection away from tallest stereocilia = tip link relaxation = channel closing = hyperpolarization
Step 2 - Exit from hair cells (basolateral surface):
- K+ exits the hair cell basolaterally into the perilymphatic space
- Key channel: KCNQ4 (a K+ channel on the basolateral membrane of hair cells, especially outer hair cells)
- Loss-of-function mutations in KCNQ4 cause progressive nonsyndromic deafness (DFNA2)
Step 3 - Return to spiral ligament (two pathways):
Pathway A - Epithelial gap junction network:
- K+ enters supporting cells (Deiters, Hensen, Boettcher, Claudius cells) through gap junctions
- Travels intracellularly through the epithelial gap junction network to the spiral ligament
- Key proteins: Connexin 26 (CX26/GJB2) and Connexin 30 (CX30/GJB6)
Pathway B - Perilymphatic diffusion:
- K+ diffuses extracellularly via the perilymphatic space toward the spiral ligament
Step 4 - K+ uptake by spiral ligament fibrocytes:
- Type II fibrocytes in the spiral ligament take up K+ and transfer it to type I fibrocytes
- This is mediated via the connective tissue gap junction network (also connexin-dependent)
- K+ travels intracellularly from fibrocytes to basal cells of stria vascularis
Step 5 - Entry into stria vascularis (basal cell layer):
- K+ enters basal and intermediate cells through gap junctions
- Intermediate cells release K+ into the intrastrial space via KCNJ10 channels (Kir4.1)
- KCNJ10 mutations cause EAST/SeSAME syndrome (epilepsy, ataxia, sensorineural deafness, tubulopathy)
- The intrastrial potential is ~+100 mV
Step 6 - Uptake by marginal cells from intrastrial space:
- Marginal cells actively take up K+ from the intrastrial compartment using two mechanisms:
- NKCC1 cotransporter (Na+/K+/2Cl- cotransporter) - brings Na+, K+, and 2Cl- in together, driven by the Na+ gradient
- Na+/K+-ATPase - pumps Na+ out into intrastrial space and K+ into marginal cell; creates the Na+ gradient that drives NKCC1
- Na+ is pumped back into intrastrial space; Cl- exits basolaterally via ClC-K/Barttin channels
Step 7 - K+ secretion into endolymph (apical surface of marginal cells):
- K+ is secreted from marginal cells into the scala media through KCNQ1/KCNE1 K+ channels
- Driven by the high positive resting potential of marginal cells
- This final step replenishes the endolymph with high K+ and maintains the ECP
VII. Ion Channels and Transporters - Summary Table
| Channel/Transporter | Location | Function | Disease if Mutated |
|---|---|---|---|
| TMC1/TMC2 | Hair cell stereociliary tips | Mechanotransduction - K+ entry | Nonsyndromic deafness |
| KCNQ4 | Hair cell basolateral membrane | K+ exit from hair cell | DFNA2 (progressive SNHL) |
| Connexin 26 (GJB2) | Epithelial gap junctions | K+ recycling via supporting cells | Most common cause of nonsyndromic hereditary HL |
| Connexin 30 (GJB6) | Epithelial gap junctions | K+ recycling | DFNA3, DFNB1 |
| Connexin 31, 43 | Gap junctions | K+ recycling | Nonsyndromic HL |
| KCNJ10 (Kir4.1) | Intermediate cells of stria vascularis | K+ release into intrastrial space; generates ECP | EAST/SeSAME syndrome |
| NKCC1 | Marginal cells (basolateral) | Na+/K+/2Cl- uptake into marginal cell | Target of loop diuretics (furosemide) - ototoxicity |
| Na+/K+-ATPase | Marginal cells (basolateral) | Creates Na+ gradient; K+ uptake | Blocked by ouabain; aminoglycoside toxicity |
| KCNQ1/KCNE1 | Marginal cells (apical) | K+ secretion into endolymph | Jervell and Lange-Nielsen syndrome (HL + cardiac arrhythmia) |
| ClC-K/Barttin | Marginal cells (basolateral) | Cl- recycling from marginal cell | Bartter syndrome type 4 (deafness + renal salt wasting) |
VIII. Calcium Homeostasis in Endolymph
Endolymph has an unusually low Ca2+ concentration (~20 µM), far below normal extracellular levels (~1.2 mM). This is physiologically important:
- Too low Ca2+: tip links break, leading to loss of transduction
- Too high Ca2+: mechanotransduction channels are blocked
- Ca2+ also carries part of the transduction current and regulates adaptation via myosin-1c motor activity
- Ca2+-ATPases, Na+/Ca2+ exchangers, and Ca2+-permeable channels in cochlear cell membranes regulate this concentration
IX. Volume Regulation of Endolymph
Two models of endolymph fluid dynamics have been proposed:
-
Longitudinal flow model: Endolymph is secreted by the stria vascularis and flows longitudinally through the cochlear duct to be reabsorbed in the endolymphatic sac. Disruption of this flow (endolymphatic sac dysfunction) leads to endolymphatic hydrops (Meniere's disease).
-
Radial flow model: Local secretion and reabsorption occurs radially - stria vascularis secretes; outer sulcus cells and Reissner's membrane cells reabsorb Na+ (maintaining low Na+ of endolymph).
Under pathological conditions, failure of volume regulation leads to endolymphatic hydrops - the pathological basis of Meniere's disease.
X. Clinical Correlations
A. Hereditary Hearing Loss (Connexin Mutations)
- GJB2 (Connexin 26) mutations account for ~50% of nonsyndromic recessive deafness (DFNB1)
- Impairs K+ recycling through the epithelial gap junction network
- Also expressed in stria vascularis; GJB2 mutations disrupt multiple steps of K+ circulation
B. Jervell and Lange-Nielsen (JLN) Syndrome
- Mutations in KCNQ1 or KCNE1 genes
- Prevents K+ secretion from marginal cells into endolymph
- Features: profound congenital SNHL + prolonged QT interval + risk of sudden cardiac death
- Autosomal recessive
C. Loop Diuretic Ototoxicity (Furosemide)
- Furosemide inhibits NKCC1 in marginal cells
- Suppresses K+ secretion into endolymph
- Abolishes the ECP - reversible hearing loss (dose-dependent)
- Potentiated by aminoglycosides (synergistic ototoxicity)
D. Aminoglycoside Ototoxicity
- Aminoglycosides inhibit Na+/K+-ATPase
- Disrupts the Na+ gradient driving NKCC1
- Also cause direct hair cell toxicity (reactive oxygen species)
- Combined use with furosemide is highly ototoxic - must be avoided
E. Meniere's Disease
- Endolymphatic hydrops: excess endolymph volume
- Possible mechanism: failure of endolymphatic sac reabsorption
- Distension of Reissner's membrane and rupture events cause mixing of endo- and perilymph
- Leads to episodic SNHL, tinnitus, aural fullness, and vertigo
F. EAST/SeSAME Syndrome
- Mutation in KCNJ10 (intermediate cell K+ channel in stria vascularis)
- Features: Epilepsy, Ataxia, Sensorineural deafness, Tubulopathy
- Rare, autosomal recessive
G. Bartter Syndrome Type 4
- Mutations in Barttin (ClC-K Cl- channel subunit) in marginal cells
- Disrupts Cl- recycling, secondarily impairs K+ secretion
- Features: sensorineural deafness + severe salt-wasting nephropathy
XI. Summary Flowchart: K+ Transport Circuit
ENDOLYMPH (high K+, +85 mV)
|
| K+ enters hair cell apically via TMC1/TMC2
↓
HAIR CELL (depolarization → neurotransmission)
|
| K+ exits basolaterally via KCNQ4
↓
PERILYMPH / SUPPORTING CELLS
↙ ↘
Via epithelial Via perilymph
gap junctions extracellular
(CX26/CX30) diffusion
↘ ↙
SPIRAL LIGAMENT FIBROCYTES (Type II → Type I)
|
| Via connective tissue gap junction network
↓
BASAL & INTERMEDIATE CELLS OF STRIA VASCULARIS
|
| K+ released into intrastrial space via KCNJ10
↓
INTRASTRIAL COMPARTMENT (+100 mV)
|
| NKCC1 + Na+/K+-ATPase take K+ into MARGINAL CELLS
↓
MARGINAL CELLS
|
| K+ secreted apically into scala media via KCNQ1/KCNE1
↓
ENDOLYMPH (K+ replenished, ECP maintained)
References
- Cummings Otolaryngology Head and Neck Surgery, 7th ed. - Chapter 128: Inner Ear Physiology
- Shambaugh's Surgery of the Ear, 6th ed. - Chapter 4: Physiology of the Cochlea (Stria Vascularis, Endocochlear Potential and Potassium Homeostasis, Cochlear Fluid Homeostasis)
- Bradley & Daroff's Neurology in Clinical Practice - Normal Anatomy and Physiology of the Inner Ear
This is a shared conversation. Sign in to Orris to start your own chat.