Could you please explain everything about resting membrane potential and action potential from Stuart fox mbbs and Guyton and hall mbbs books?
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Searching the Web
Stuart Fox Human Physiology resting membrane potential action potential summary
Reading Web Page
https://houptlab.org/courses/humanphys/Ch7-2-Action%20Potent…
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Loading Image
Reading File
Finding Sources
Reading Web Page
https://quizlet.com/577924629/human-physiology-stuart-ira-fo…
Reading File
I now have all the content I need. Let me compile a thorough, well-organized response.
Resting Membrane Potential and Action Potential
A note on sources
Guyton and Hall Textbook of Medical Physiology (14th ed., Chapter 5) is in the library and is quoted directly below. Stuart Fox's Human Physiology is not in this library, but its Chapter 7 content is well-documented in open course materials from Florida State University (Dr. T. Houpt's PCB 4701 course, directly based on Fox), and the core physiology is identical across both books - differences in emphasis are noted where they exist.
PART 1 - RESTING MEMBRANE POTENTIAL (RMP)
1.1 What Is the RMP?
The resting membrane potential of large nerve fibers is approximately -70 mV (inside negative relative to outside). The membrane is described as "polarized" in this state.
"The resting membrane potential of large nerve fibers when they are not transmitting nerve signals is about −70 mV. That is, the potential inside the fiber is 70 mV more negative than the potential in the extracellular fluid on the outside of the fiber."
- Guyton and Hall, p. 78
Stuart Fox (Ch. 7) states the same value: Vm (resting) = -70 mV, with K⁺ being highly permeable at rest, and the cell being high in K⁺ inside and high in Na⁺ outside.
1.2 The Na⁺-K⁺ Pump (Electrogenic Pump)
The foundation of the RMP is the Na⁺-K⁺ ATPase pump, which continuously:
- Pumps 3 Na⁺ OUT for every 2 K⁺ IN
- Creates a net loss of positive charges from inside the cell
- Is therefore electrogenic - it directly contributes about -4 mV to the RMP
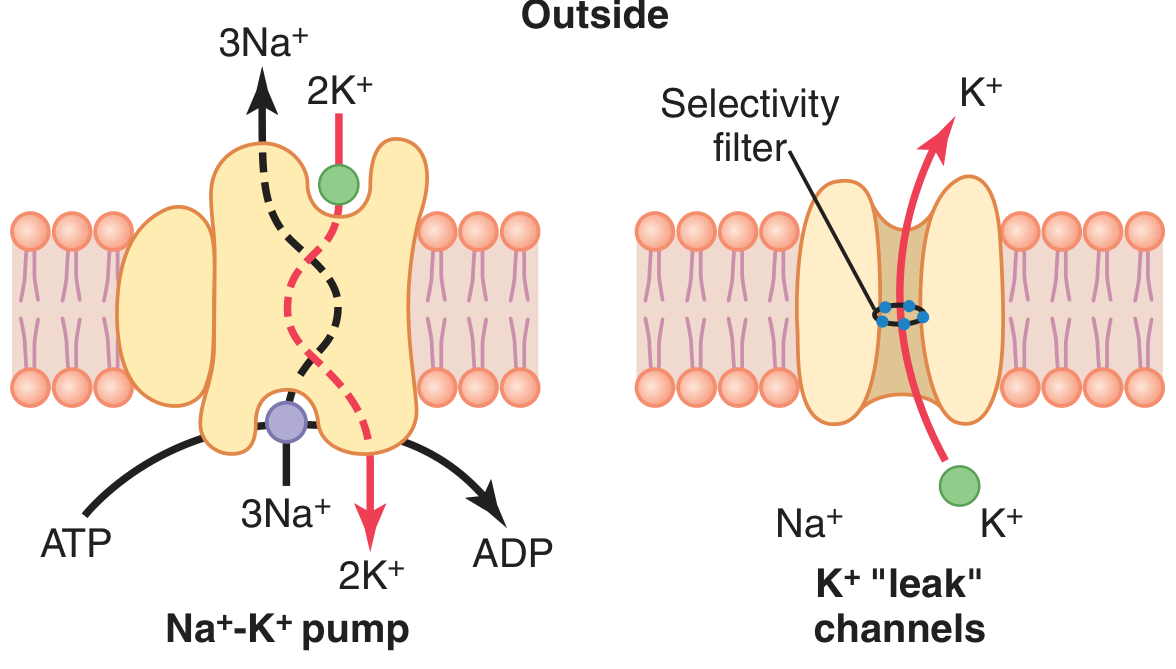
Guyton Fig. 5.4: The Na⁺-K⁺ pump (left) pumps 3 Na⁺ out and 2 K⁺ in per cycle. K⁺ "leak" channels (right) allow passive K⁺ efflux down its concentration gradient.
This pump establishes the following ion concentration gradients:
| Ion | Outside | Inside | Ratio |
|---|---|---|---|
| Na⁺ | 142 mEq/L | 14 mEq/L | Na_in/Na_out = 0.1 |
| K⁺ | 4 mEq/L | 140 mEq/L | K_in/K_out = 35.0 |
1.3 K⁺ Leak Channels and the Potassium Diffusion Potential
The membrane at rest is ~100x more permeable to K⁺ than to Na⁺, due to open K⁺ "leak" channels (tandem pore domain / K⁺ leak channels). These allow K⁺ to diffuse out down its concentration gradient, leaving negative charges behind inside.
The Nernst Equation calculates the equilibrium potential for any single ion:
If only K⁺ could cross the membrane, the Nernst potential for K⁺ (K_in/K_out = 35) = -94 mVIf only Na⁺ could cross, the Nernst potential for Na⁺ (Na_in/Na_out = 0.1) = +61 mV
1.4 The Goldman Equation - Combining All Ions
Because the membrane is permeable to both ions (though unequally), the Goldman-Hodgkin-Katz (GHK) equation is used, which weights each ion's contribution by its permeability. Since K⁺ permeability is ~100x Na⁺ permeability:
- Diffusion of K⁺ and Na⁺ together → -86 mV
- Adding the electrogenic pump contribution (−4 mV) → net ≈ −90 mV
The measured RMP in large nerve fibers is −70 mV (slightly less negative due to contributions of Cl⁻ and other ions).
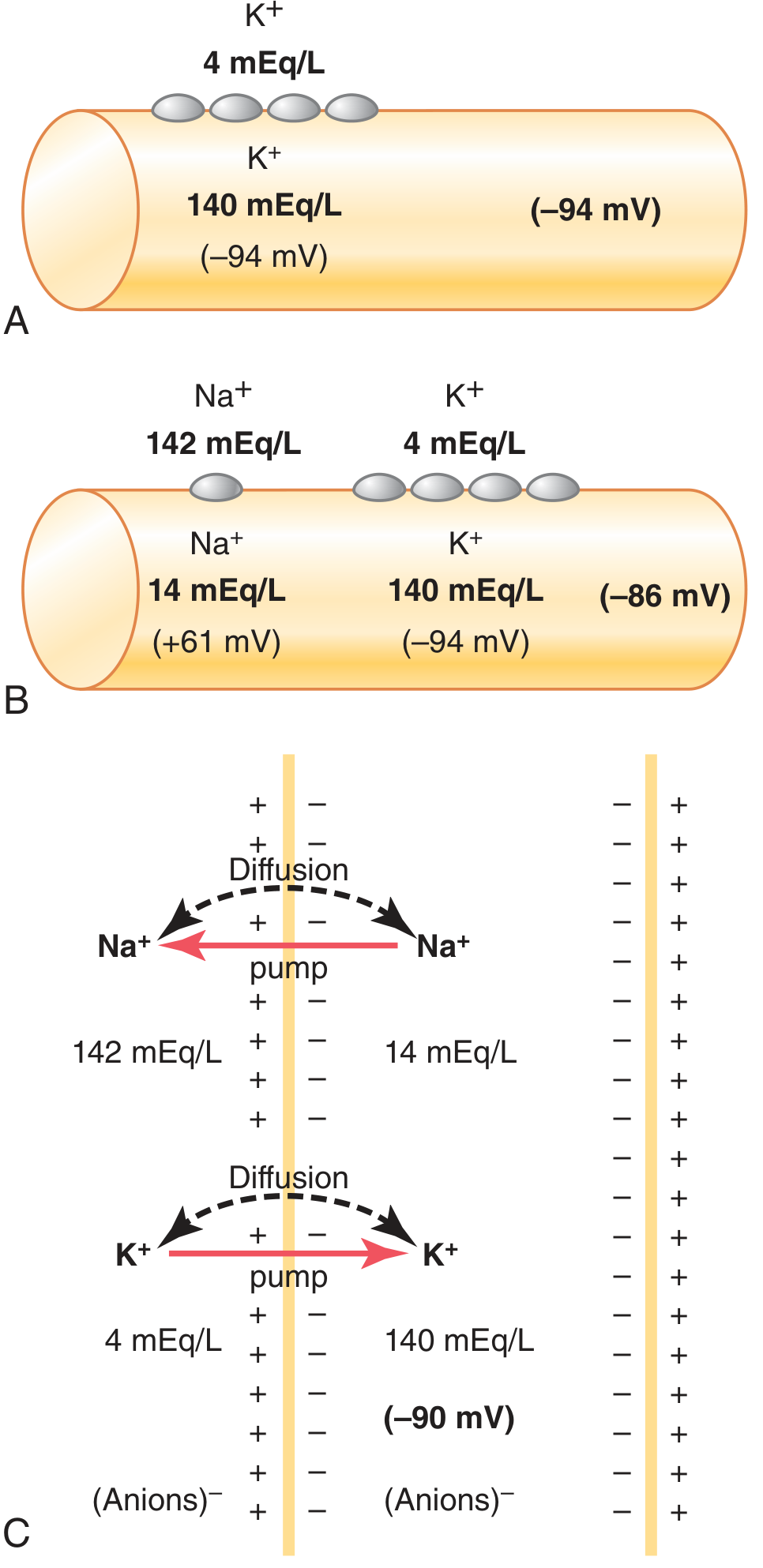
Guyton Fig. 5.5: (A) K⁺ diffusion alone → -94 mV. (B) K⁺ + Na⁺ diffusion → -86 mV. (C) Adding the Na⁺-K⁺ pump → -90 mV.
1.5 Stuart Fox's Emphasis on RMP
In Fox Ch. 7, the RMP is explained through two key concepts:
- Equilibrium potential - for each ion, there is a voltage at which the electrical force exactly balances the diffusion force (Nernst potential). K⁺ equilibrium potential ≈ -90 mV; Na⁺ equilibrium potential ≈ +60 mV.
- Relative permeability - the resting RMP of -70 mV is a compromise between these two opposing equilibrium potentials, weighted heavily toward K⁺ because of its much higher resting permeability.
"Relative membrane permeability to K⁺ is the essential factor that produces resting membrane potential." - Fox Ch. 7
PART 2 - ACTION POTENTIAL (AP)
2.1 Overview
The action potential is a rapid, transient reversal of membrane polarity that propagates along excitable membranes. It follows the all-or-nothing principle.
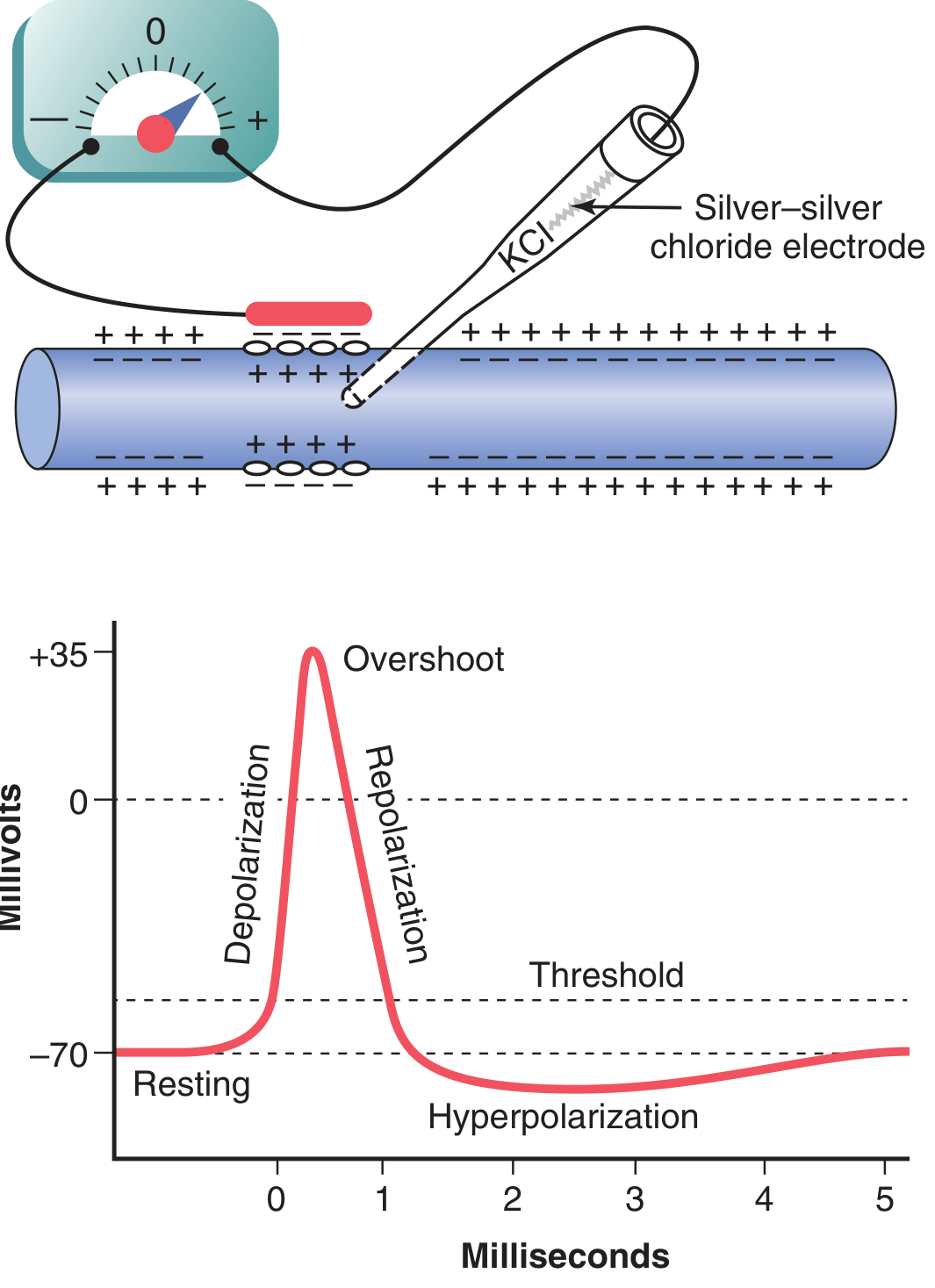
Guyton Fig. 5.6: The classic action potential waveform - showing resting stage at -70 mV, rapid depolarization, overshoot to +35 mV, repolarization, and brief hyperpolarization (undershoot) before return to rest.
2.2 Phases of the Action Potential
Phase 1 - Resting Stage
- Membrane potential = -70 mV
- Membrane is said to be "polarized"
- Voltage-gated Na⁺ channels: CLOSED (activation gate shut, inactivation gate open)
- Voltage-gated K⁺ channels: CLOSED
Phase 2 - Depolarization
- A stimulus depolarizes the membrane to the threshold (~-55 mV, or about 15 mV above resting)
- At threshold, voltage-gated Na⁺ channels snap open rapidly
- Na⁺ rushes IN along its electrochemical gradient (Na⁺ is high outside, and the inside is negative)
- Membrane potential shoots up: -70 mV → 0 mV → +35 mV (overshoot)
- This is a positive feedback cycle - Na⁺ entry makes inside more positive → more Na⁺ channels open → more Na⁺ enters
"This process is a positive-feedback cycle that, once the feedback is strong enough, continues until all the voltage-gated sodium channels have become activated (opened)."
- Guyton and Hall, p. 82
Phase 3 - Repolarization
- Within a fraction of a millisecond of opening:
- Na⁺ channels inactivate (the inactivation gate closes) - Na⁺ entry stops
- Voltage-gated K⁺ channels open (delayed, slower response)
- K⁺ rushes OUT, restoring the negative charge inside
- Membrane returns from +35 mV back toward -70 mV
Phase 4 - Hyperpolarization (Undershoot / After-Hyperpolarization)
- K⁺ channels stay open slightly longer than needed → membrane overshoots past -70 mV → becomes briefly more negative (e.g., -75 to -80 mV)
- As K⁺ channels close, membrane returns to normal resting -70 mV
2.3 Voltage-Gated Channels: The Molecular Machinery
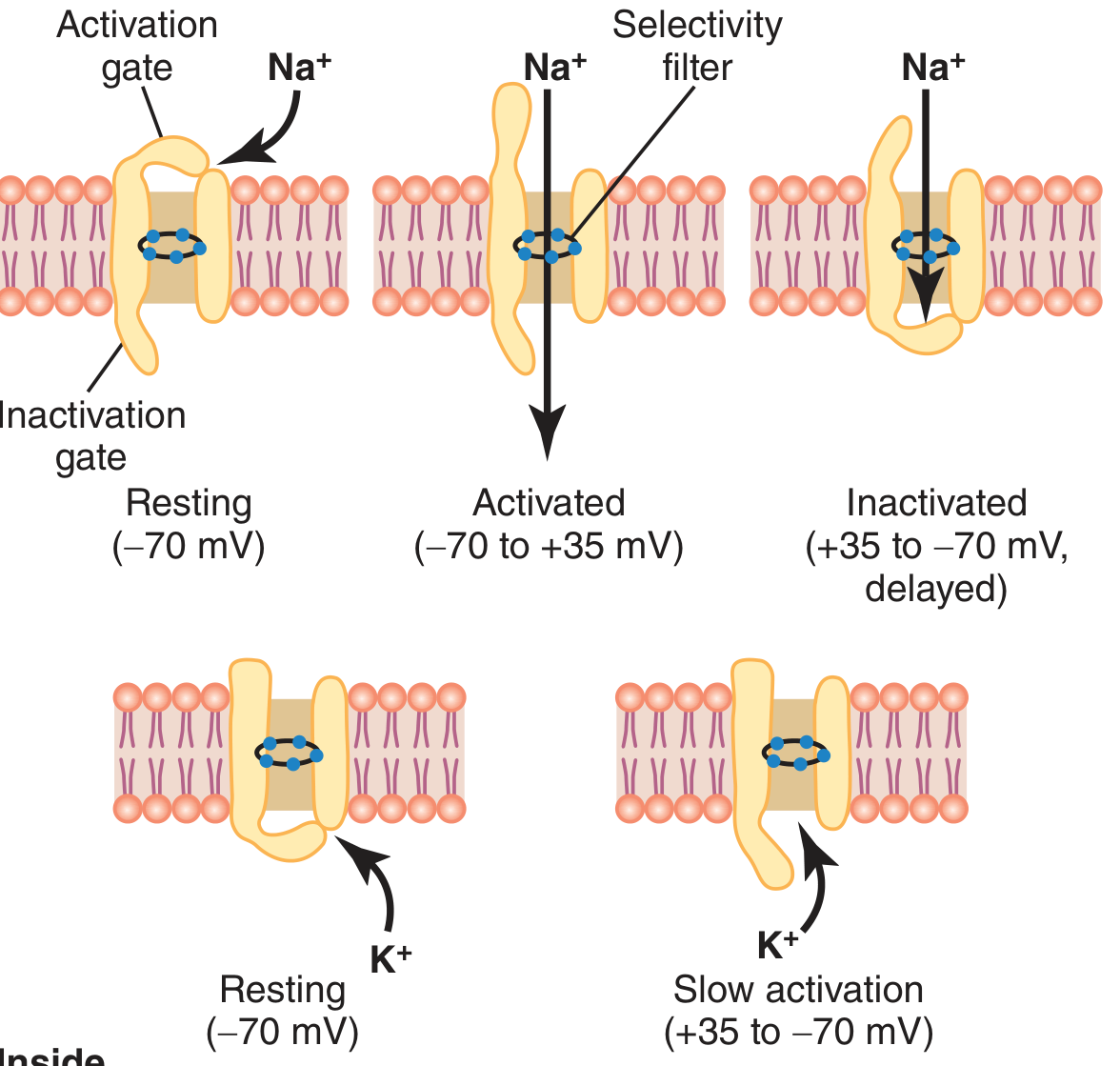
Guyton Fig. 5.7 - Na⁺ channel (top row) has TWO gates: activation gate (m-gate, outer) and inactivation gate (h-gate, inner). K⁺ channel (bottom) has one gate that opens slowly.
Voltage-Gated Na⁺ Channel - Three States:
| State | Activation Gate | Inactivation Gate | Occurs When |
|---|---|---|---|
| Resting (closed) | Closed | Open | At -70 mV |
| Activated (open) | Open | Open | At threshold, during depolarization |
| Inactivated (closed) | Open | Closed | Peak of AP; channel cannot reopen until membrane repolarizes |
"The same increase in voltage that opens the activation gate also closes the inactivation gate. The inactivation gate, however, closes a few 10,000ths of a second after the activation gate opens."
- Guyton and Hall, p. 81
Key fact: Na⁺ channel permeability increases 500 to 5,000-fold when activated.
Voltage-Gated K⁺ Channel:
- Has one gate; opens slowly during depolarization (takes ~10x longer than Na⁺ channel)
- Opens at the peak of the AP, driving repolarization
- Closes slowly → causes the hyperpolarization undershoot
Stuart Fox adds - Sodium Conductance vs. Potassium Conductance:
Fox emphasizes "conductance" (=permeability) changes as the key to understanding the AP: Na⁺ conductance rises sharply first, then falls; K⁺ conductance rises later and falls gradually - the AP waveform is the direct consequence of these two overlapping conductance changes (Fox Fig. 7.19).
2.4 Threshold and the All-or-Nothing Principle
- Threshold is reached when the inward Na⁺ current exceeds the outward K⁺ current
- A depolarization of 15-30 mV from rest is typically needed (from -70 mV up to about -55 mV)
- Once threshold is crossed, a full AP fires regardless of stimulus strength - this is the all-or-nothing principle
- Subthreshold stimuli cause only local, graded potentials that fade without propagating
2.5 Propagation of the Action Potential
Once an AP fires at one point on the membrane:
- The depolarized region creates local circuits of ionic current flow - positive ions flow inward through the depolarized zone, then spread along the axon core
- These currents depolarize adjacent membrane segments to threshold
- New APs fire in adjacent regions
- The wave of depolarization travels down the entire length of the fiber
"The arrows show a local circuit of current flow from the depolarized areas of the membrane to the adjacent resting membrane areas...the sodium channels in these new areas immediately open...and the explosive action potential spreads."
- Guyton and Hall, p. 84
Direction: The AP spreads in all directions away from the stimulus.
Fox's description: The AP is a "wave of depolarization spreading down the axon with voltage-sensitive Na⁺ channels opening" - it is ions moving IN and OUT, not current traveling down the axon length.
2.6 Myelination and Saltatory Conduction
In myelinated nerve fibers, the myelin sheath acts as an insulator, and ionic exchange can only occur at the nodes of Ranvier. The AP "jumps" from node to node - called saltatory conduction - making propagation:
- Much faster (up to 120 m/s in large myelinated fibers vs. 0.5 m/s in unmyelinated)
- Energetically efficient (Na⁺/K⁺ pumps only need to restore ions at the nodes)
2.7 Refractory Periods
| Period | Duration (large myelinated nerve) | Mechanism | Can fire again? |
|---|---|---|---|
| Absolute refractory period | ~1 ms | Na⁺ channels are inactivated; inactivation gates CANNOT reopen until membrane repolarizes | No, even with maximal stimulus |
| Relative refractory period | 2-4 ms (during hyperpolarization) | Na⁺ channels recovering; K⁺ channels still open; threshold elevated | Yes, but requires a STRONGER stimulus |
"The period during which a second action potential cannot be elicited, even with a strong stimulus, is called the absolute refractory period...about 1 millisecond. Therefore, one can readily calculate that such a fiber can transmit a maximum of about 1000 impulses per second."
- Guyton and Hall, p. 84
Functional significance: Refractory periods:
- Ensure APs travel in one direction only (the region behind the AP is in absolute refractory period)
- Set a maximum firing frequency (~1000/sec for large myelinated fibers)
2.8 Calcium's Role (Guyton)
- Normal extracellular Ca²⁺ stabilizes the sodium channel by binding to the exterior and raising the threshold for Na⁺ channel activation
- Hypocalcemia (low Ca²⁺) → reduced threshold → spontaneous nerve firing → tetany
- Hypercalcemia → elevated threshold → reduced excitability
- Voltage-gated Ca²⁺ channels (slow channels) also contribute to APs in cardiac and smooth muscle
2.9 Energy Metabolism After Action Potentials
Each AP moves a tiny number of ions across the membrane (essentially unmeasurable after a single AP). However, the Na⁺-K⁺ pump must eventually restore gradients. Pump activity is stimulated by rising intracellular Na⁺ (approximately proportional to the cube of intracellular Na⁺ concentration). A nerve fiber can fire 100,000 to 50 million impulses before the gradient fails without pumping.
COMPARISON: Guyton & Hall vs. Stuart Fox
| Feature | Guyton & Hall | Stuart Fox (Ch. 7) |
|---|---|---|
| RMP value | -70 mV | -70 mV |
| RMP mechanism | Na⁺-K⁺ pump + K⁺ leak channels + Goldman equation | Equilibrium potentials (Nernst) + relative permeability |
| Threshold | ~-55 mV (~15-30 mV depolarization) | Threshold = level where AP becomes self-sustaining |
| AP overshoot | +35 mV | +50 mV (approaches ENa) |
| Channel gating | Activation gate + inactivation gate | Voltage-sensitive Na⁺ and K⁺ channels; conductance changes |
| Propagation | Local circuits of current flow | Wave of depolarization; ions move in/out (not down the axon) |
| Absolute refractory | ~1 ms | Na⁺ channels inactivated |
| Relative refractory | 2-4 ms | K⁺ channels still open, threshold raised |
| Emphasis | Quantitative (Nernst, Goldman, ion concentrations, mV values) | Conceptual clarity with conductance-based model (Fox Fig. 7.19) |
Summary Flow (Both Books)
RMP (-70 mV)
│
│ ← Stimulus (depolarization to threshold ~-55 mV)
▼
Positive feedback: Na⁺ channels open → Na⁺ rushes in → more channels open
│
▼
Depolarization → Overshoot (+35 mV)
│
▼
Na⁺ channels inactivate → K⁺ channels open → K⁺ rushes out
│
▼
Repolarization → Hyperpolarization (undershoot)
│
▼
K⁺ channels close → Return to RMP (-70 mV)
│
▼
Na⁺-K⁺ pump restores concentration gradients (slow, metabolic)
Key Sources:
- All quoted content and diagrams: Guyton and Hall Textbook of Medical Physiology, 14th ed., Chapter 5 (pp. 77-89)
- Stuart Fox Ch. 7 content: PCB 4701 Action Potential lecture materials, based on Fox's Human Physiology, 14th ed.
This is a shared conversation. Sign in to Orris to start your own chat.