median nerve anatomy
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Finding Sources
Reading File
Reading File
Loading Image
I now have all the information needed for a thorough answer. Here is the full breakdown:
Median Nerve Anatomy
Origin and Roots
The median nerve arises from the lateral (C6, C7) and medial (C8, T1) cords of the brachial plexus. Its two roots unite anterior to the third part of the axillary artery to form the nerve proper. The full spinal root contribution is C5-C6-C7 (via lateral cord) and C8-T1 (via medial cord), making it a C6-T1 nerve overall.
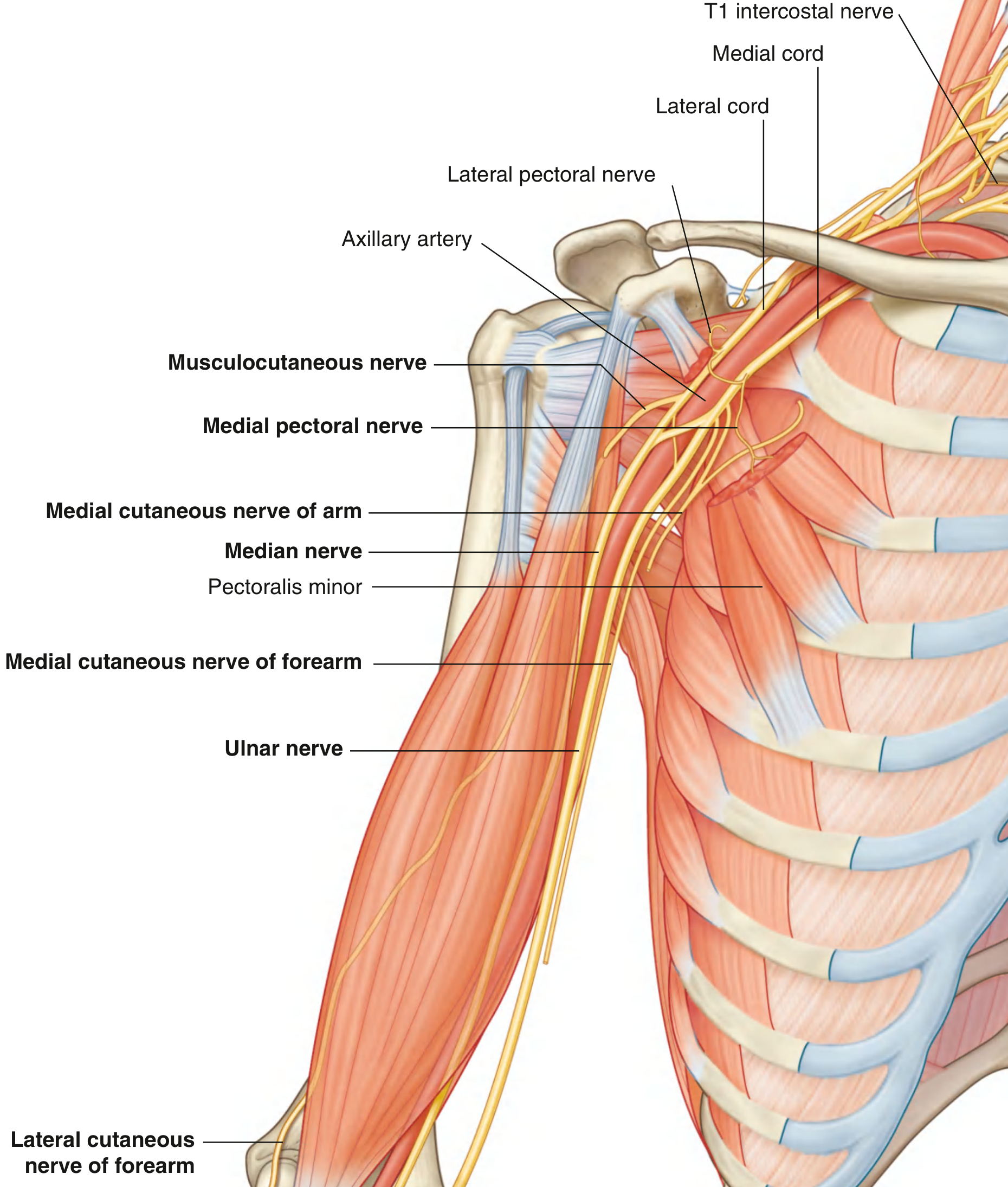
Course Through the Arm
- The nerve enters the arm from the axilla at the inferior margin of teres major and descends in the anterior compartment, closely related to the brachial artery throughout.
- Proximally: lies lateral to the brachial artery.
- Distally: crosses to the medial side of the brachial artery (in 74% of cases, the crossing is in the lower third of the arm) and lies anterior to the elbow joint.
- No major branches are given off in the arm - though a small branch to the pronator teres may arise just proximal to the elbow.
Course Through the Forearm
The nerve leaves the cubital fossa by passing between the two heads of the pronator teres, then passes between the humero-ulnar and radial heads of flexor digitorum superficialis.
It then travels distally deep to (on the deep surface of) flexor digitorum superficialis. Just proximal to the wrist it becomes more superficial, lying between the tendons of palmaris longus and flexor carpi radialis, before entering the carpal tunnel.
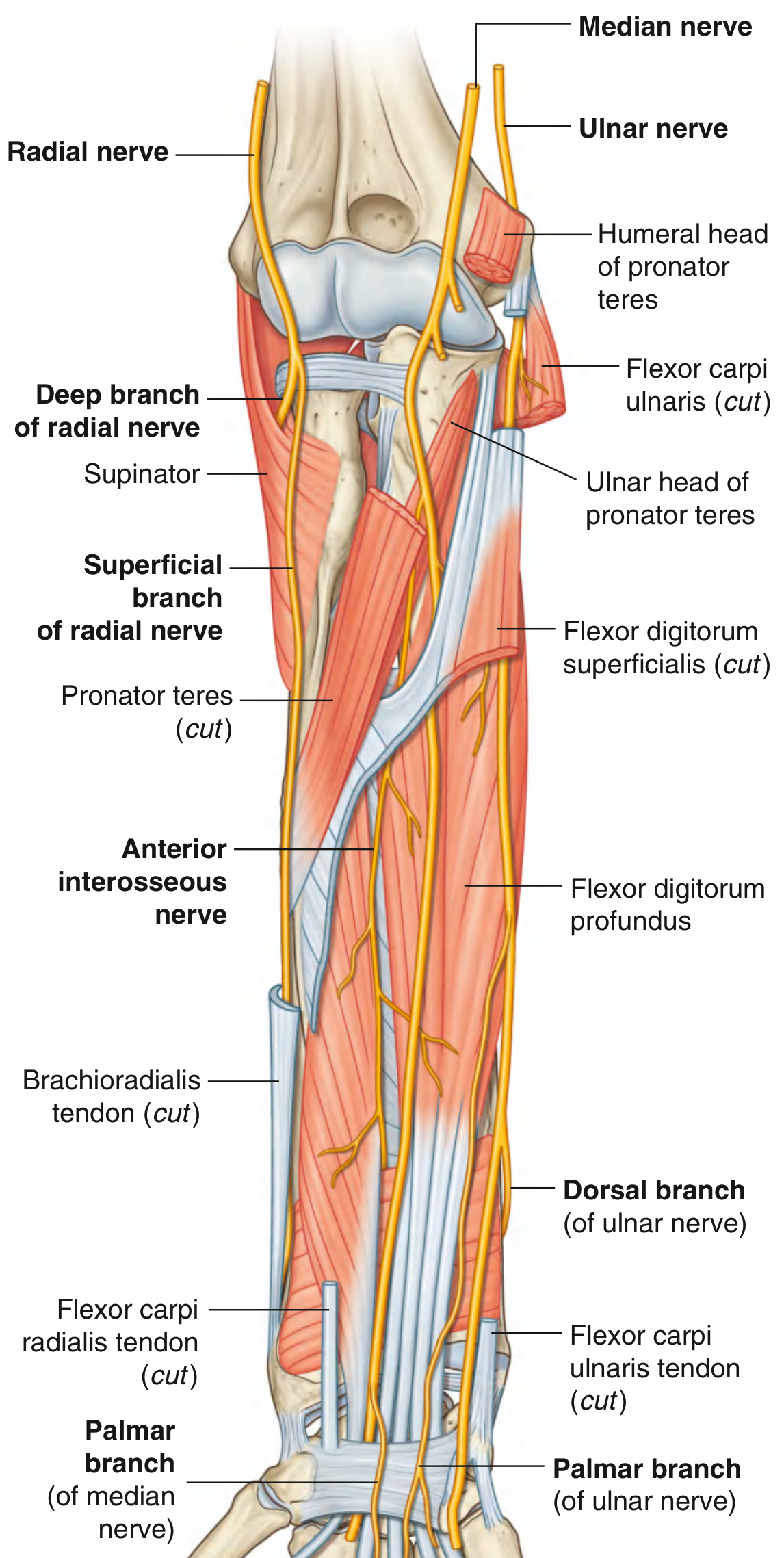
Branches in the Forearm
| Branch | Origin | Muscles Innervated |
|---|---|---|
| Muscular branches | Just distal to elbow | Pronator teres, flexor carpi radialis, palmaris longus, flexor digitorum superficialis |
| Anterior interosseous nerve (AIN) | Between the two heads of pronator teres | Flexor pollicis longus, lateral half of flexor digitorum profundus (index + middle), pronator quadratus |
| Palmar cutaneous branch | Just proximal to flexor retinaculum (passes superficial to it) | Skin over the base of the palm and thenar eminence |
The AIN is the largest branch of the median nerve in the forearm - it is a pure motor nerve, descending with the anterior interosseous artery on the anterior surface of the interosseous membrane. It terminates in articular branches to the distal forearm and wrist joints.
The palmar cutaneous branch is a key anatomical point: it travels subcutaneously, bypassing the carpal tunnel - so it is spared in carpal tunnel syndrome, which preserves sensation over the thenar eminence.
Course Through the Wrist and Hand
The median nerve enters the palm through the carpal tunnel, passing deep to the flexor retinaculum alongside the nine flexor tendons (4 from FDS, 4 from FDP, 1 from FPL). At the distal margin of the retinaculum, it divides into:
1. Recurrent (Thenar) Branch
- Arises from the lateral side of the median nerve just distal to the flexor retinaculum.
- Curves back proximally over the flexor retinaculum.
- Innervates the three thenar muscles: abductor pollicis brevis, flexor pollicis brevis (superficial head), and opponens pollicis.
- Responsible for opposition of the thumb.
2. Palmar Digital Branches
- Pass deep to the palmar aponeurosis and superficial palmar arch.
- Provide sensation to the palmar surface of the lateral 3.5 digits (thumb, index, middle, lateral half of ring finger), plus the nail beds (dorsal distal phalanges) of the same digits.
- Also innervate the lateral two lumbrical muscles (1st and 2nd lumbricals).
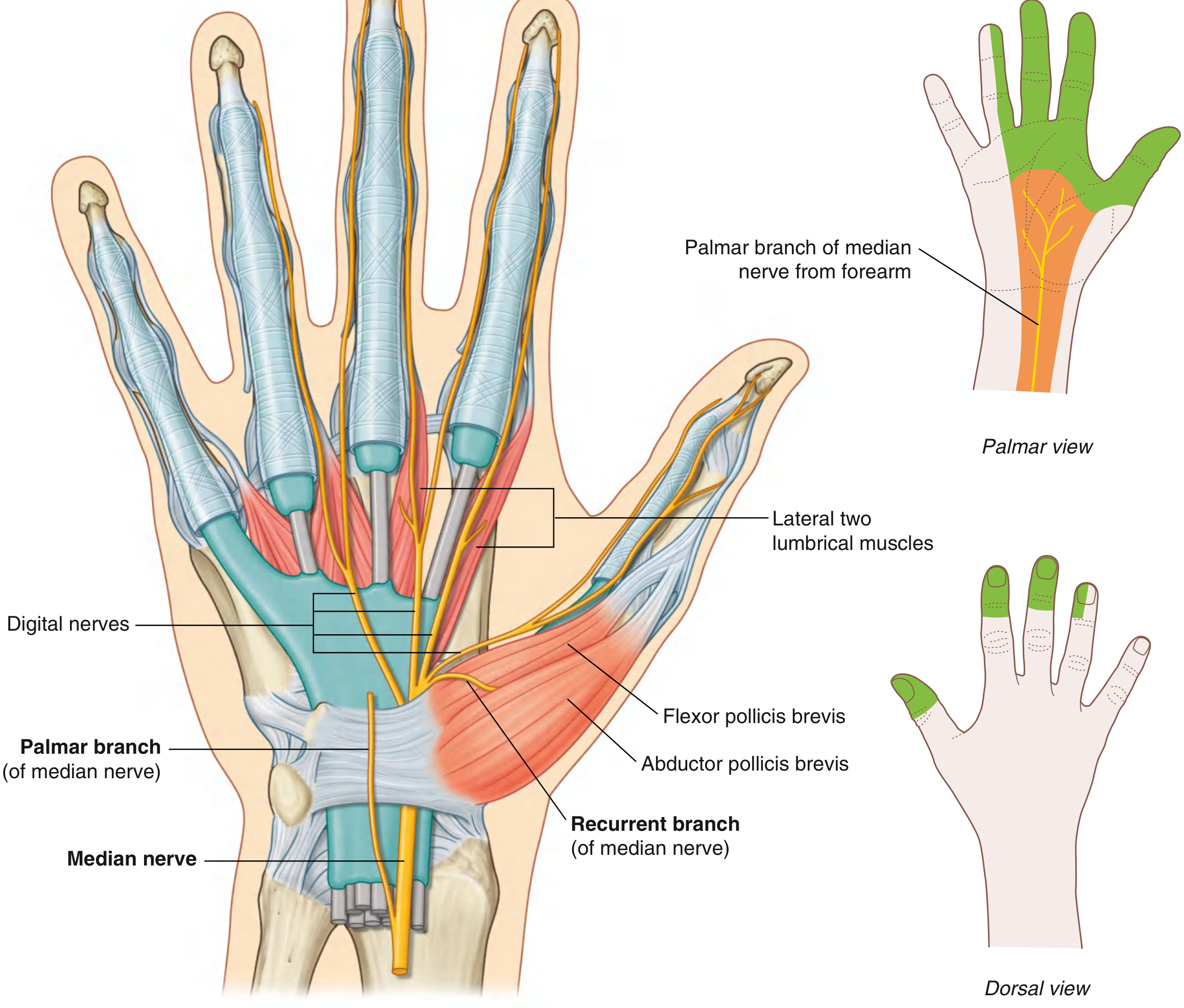
Motor Summary
| Region | Muscles |
|---|---|
| Forearm (anterior compartment, mostly superficial/intermediate) | Pronator teres, FCR, palmaris longus, FDS |
| Forearm (deep layer, via AIN) | FPL, lateral FDP (index + middle fingers), pronator quadratus |
| Thenar eminence (hand) | Abductor pollicis brevis, opponens pollicis, flexor pollicis brevis (superficial head) |
| Hand (intrinsics) | Lateral two lumbricals (1st and 2nd) |
Not innervated by median nerve: FCU, medial half of FDP (ring and little fingers) - these belong to the ulnar nerve.
Sensory Summary
- Palm: base and central palm via the palmar cutaneous branch (spared in CTS)
- Digits (palmar): thumb, index, middle, and lateral half of ring finger
- Digits (dorsal): distal phalanges (nail beds) of the same 3.5 fingers
Key Clinical Correlations
| Injury Level | Deficit |
|---|---|
| High lesion (elbow/above) | Loss of pronation, wrist flexion (radial deviation preserved), loss of thumb IP flexion + index DIP flexion, loss of thenar function, "hand of benediction" when trying to grip (index + middle cannot flex) |
| Anterior interosseous nerve | Cannot flex thumb IP joint + index DIP ("OK sign" failure); normal sensation |
| Carpal tunnel syndrome (wrist) | Nocturnal paresthesias in lateral 3.5 fingers, thenar atrophy, weak abductor pollicis brevis; thenar skin sensation spared (palmar cutaneous branch bypasses tunnel) |
Sources: Gray's Anatomy for Students (9780323934237); Bradley and Daroff's Neurology in Clinical Practice (9780323642613); General Anatomy and Musculoskeletal System - THIEME Atlas (9781626237186)
electron transport chain
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
I have everything needed for a comprehensive answer.
Electron Transport Chain (ETC)
Overview
The ETC is the final common pathway for oxidation of fuels in the body. Reduced coenzymes (NADH and FADH₂) generated by glycolysis, the TCA cycle, and fatty acid beta-oxidation donate their electrons to a series of protein complexes embedded in the inner mitochondrial membrane. As electrons flow down the chain to oxygen, the released energy pumps H⁺ ions across the membrane, building an electrochemical gradient that drives ATP synthesis. This coupling of electron flow to ATP production is called oxidative phosphorylation (OXPHOS).
Location and Structure
The mitochondrion has two membranes:
- Outer membrane: permeable to small ions and molecules via the protein porin
- Inner membrane: highly impermeable - no H⁺, ATP, ADP, or pyruvate can cross without specific transporters. It is folded into cristae to maximize surface area. This is where the ETC complexes live.
- Matrix: gel-like interior containing TCA cycle enzymes, fatty acid oxidation enzymes, mtDNA, and the NAD⁺/FAD required as electron acceptors.
The Four Complexes
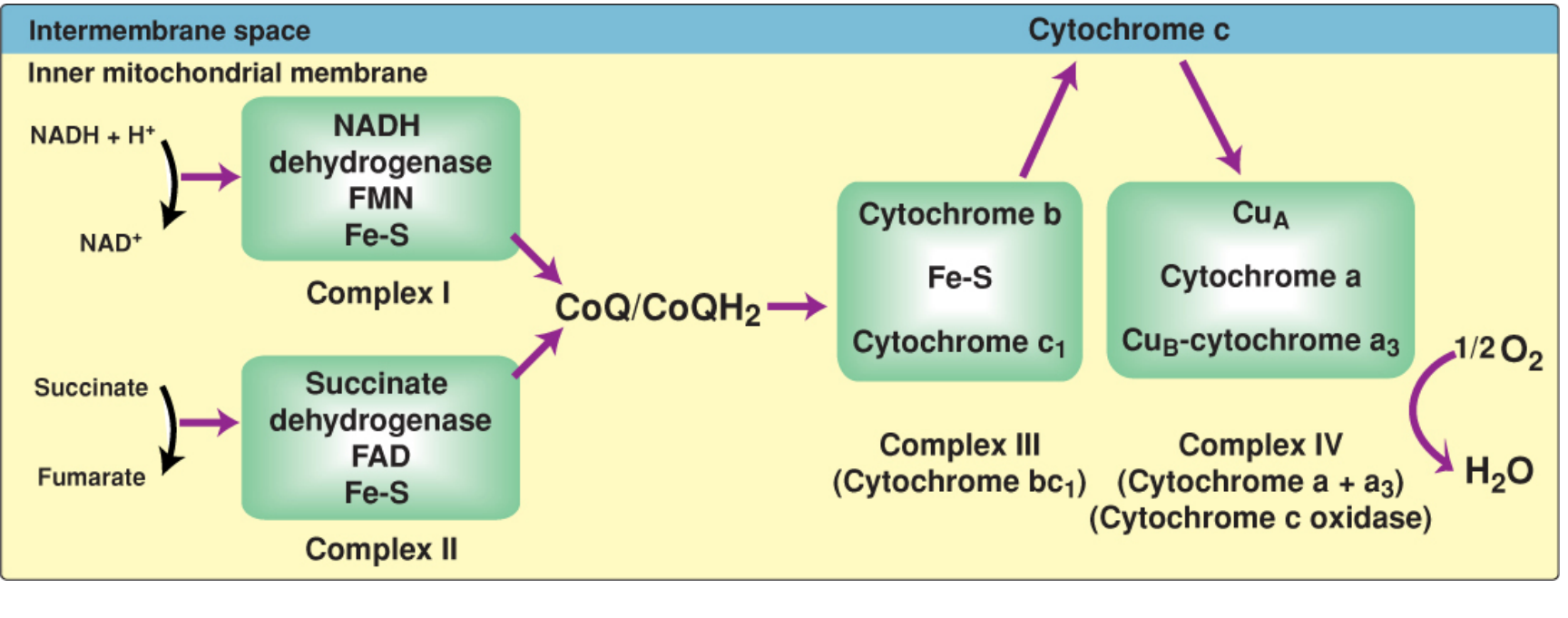
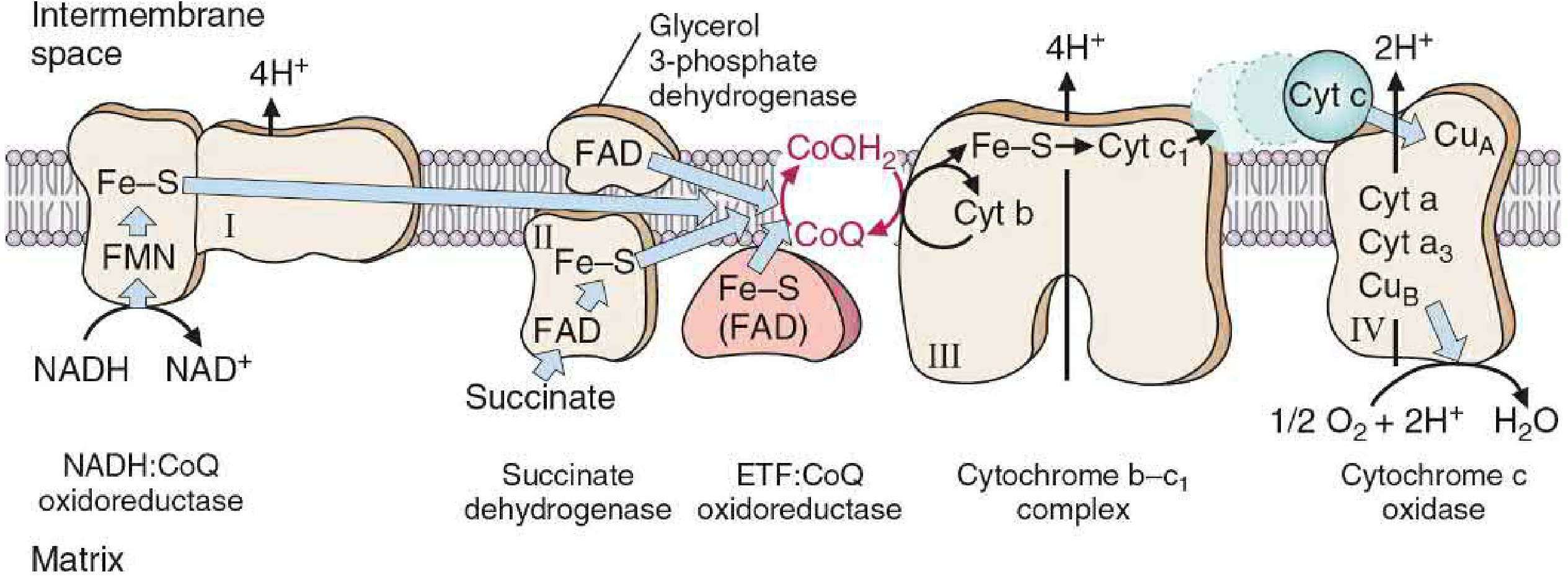
Complex I - NADH Dehydrogenase (NADH:CoQ Oxidoreductase)
- Accepts electrons from: NADH (generated by TCA cycle, pyruvate dehydrogenase, beta-oxidation)
- Prosthetic groups: FMN (flavin mononucleotide) + multiple Fe-S centers
- Electron flow: NADH → FMN → Fe-S centers → CoQ
- Protons pumped: 4 H⁺ from matrix to intermembrane space
- A massive 45-subunit complex spanning the inner membrane
Complex II - Succinate Dehydrogenase
- Accepts electrons from: FADH₂ (produced when succinate is oxidized to fumarate in the TCA cycle)
- Prosthetic groups: FAD + Fe-S centers
- Electron flow: FADH₂ → Fe-S → CoQ
- Protons pumped: 0 - this complex does not span the membrane and has no proton-pumping mechanism, which is why FADH₂ yields less ATP than NADH
- Also accepts electrons from glycerol-3-phosphate dehydrogenase and acyl-CoA dehydrogenases (fatty acid oxidation)
Mobile Carrier - Coenzyme Q (CoQ / Ubiquinone)
- A lipid-soluble quinone with a long hydrophobic isoprenoid tail - freely mobile in the lipid bilayer
- Acts as a junction point: collects electrons from Complexes I, II, glycerol-3-phosphate dehydrogenase, and ETF:CoQ oxidoreductase (fatty acid beta-oxidation)
- Transfers electrons to Complex III
- When reduced: CoQH₂ (ubiquinol)
Complex III - Cytochrome bc₁ (Ubiquinol:Cytochrome c Oxidoreductase)
- Accepts electrons from: CoQH₂
- Components: Cytochrome b, Fe-S protein (Rieske protein), cytochrome c₁
- Electron flow: CoQH₂ → cyt b → Fe-S → cyt c₁ → cytochrome c
- Protons pumped: 4 H⁺ into intermembrane space (via the Q cycle mechanism)
Mobile Carrier - Cytochrome c
- A small, water-soluble heme-protein located in the intermembrane space, loosely bound to the outer face of the inner membrane
- Shuttles single electrons from Complex III to Complex IV
- Iron in heme oscillates between Fe²⁺ and Fe³⁺
- Plays a separate role in apoptosis when released into the cytosol
Complex IV - Cytochrome c Oxidase (Cytochrome a + a₃)
- Accepts electrons from: cytochrome c
- Components: CuA, cytochrome a, cytochrome a₃, CuB
- Electron flow: cyt c → CuA → cyt a → cyt a₃ (with CuB) → O₂
- Terminal reaction: 4 electrons + O₂ + 4H⁺ → 2 H₂O
- This is the only complex where heme iron has a free coordination site that can directly bind O₂
- Protons pumped: 2 H⁺ into intermembrane space
- Requires copper (Cu) for the reaction to occur
Chemiosmosis and ATP Synthesis
The combined pumping of H⁺ at Complexes I, III, and IV creates:
- A chemical gradient (high [H⁺] in intermembrane space vs. matrix)
- An electrical gradient (positive outside, negative inside)
Together these form the proton-motive force (PMF). H⁺ ions flow back down this gradient through ATP synthase (Complex V), which uses the energy of proton flow to phosphorylate ADP → ATP. This is the binding change mechanism - the rotation of the γ-subunit driven by proton influx changes the conformation of the three αβ-subunit pairs, cycling through open, loose, and tight conformations to bind substrates, form ATP, and release the product.
ATP Yield
| Electron Donor | Protons Pumped (I+III+IV) | ATPs Produced |
|---|---|---|
| NADH | 4 + 4 + 2 = 10 H⁺ | ~2.5 ATP |
| FADH₂ (via Complex II) | 0 + 4 + 2 = 6 H⁺ | ~1.5 ATP |
- Overall free energy: NADH oxidation releases ~53 kcal/mol; FADH₂ ~41 kcal/mol
- Only ~30% of this energy is trapped as ATP; the rest is released as heat (the ETC is also the body's primary heat source)
- Approximately 3 H⁺ are needed per ATP synthesized, plus 1 H⁺ for phosphate transport into the matrix
Regulation
The ETC is regulated primarily through coupling to ATP demand:
- When cells use ATP, ADP + Pᵢ levels rise
- ADP binds ATP synthase → increased H⁺ influx → gradient dissipates
- Reduced gradient → Complexes I, III, IV pump faster → electron flow increases → O₂ consumption rises
- Increased NADH consumption signals the TCA cycle and fuel oxidation pathways to accelerate
This is why exercise increases both O₂ consumption and heat production proportionally.
Inhibitors
| Inhibitor | Site | Mechanism |
|---|---|---|
| Rotenone, Amytal | Complex I | Blocks NADH → CoQ electron transfer |
| Malonate | Complex II | Competitive inhibitor of succinate dehydrogenase |
| Antimycin A | Complex III | Blocks electron flow from cyt b to cyt c₁ |
| Cyanide (CN⁻), CO, H₂S, azide | Complex IV | Binds Fe in cyt a₃, blocks O₂ reduction |
| Oligomycin | ATP synthase | Blocks H⁺ channel (Fo subunit), halts ATP synthesis and secondarily halts the ETC |
| DNP (dinitrophenol), FCCP | Inner membrane | Proton ionophores - uncouplers: allow H⁺ to leak back without making ATP, causing heat generation and increased O₂ consumption with no ATP production |
Reactive Oxygen Species (ROS)
Electron leakage from the ETC (mainly at Complex I and III) partially reduces O₂ to superoxide (O₂⁻) rather than fully to H₂O. This generates:
- Superoxide (O₂⁻) → H₂O₂ (by superoxide dismutase, SOD) → OH• (hydroxyl radical via Fenton reaction)
Cellular defenses: SOD, catalase, glutathione peroxidase. Chronic ROS production contributes to aging, cancer, and neurodegeneration.
Cytoplasmic NADH: The Shuttles
NADH produced in the cytosol (e.g., during glycolysis) cannot directly cross the inner mitochondrial membrane. Two shuttles transfer its electrons:
- Malate-aspartate shuttle (heart, liver): delivers electrons to mitochondrial NAD⁺ → NADH → yield ~2.5 ATP
- Glycerol 3-phosphate shuttle (brain, muscle): delivers electrons to FAD → FADH₂ → yield ~1.5 ATP
Sources: Lippincott's Illustrated Reviews: Biochemistry, 8th ed. (9781975155063); Basic Medical Biochemistry: A Clinical Approach, 6e (9781975150143)
This is a shared conversation. Sign in to Orris to start your own chat.