Lac operon
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Here is a complete overview of the Lac Operon drawn from Harper's Illustrated Biochemistry (32nd Ed.) and Basic Medical Biochemistry (6th Ed.).
The Lac Operon
The lac operon of E. coli is the textbook model for coordinate prokaryotic gene regulation. It controls three enzymes for lactose catabolism using both negative and positive regulatory mechanisms.
1. Organization of the Lac Operon (~6 kb)
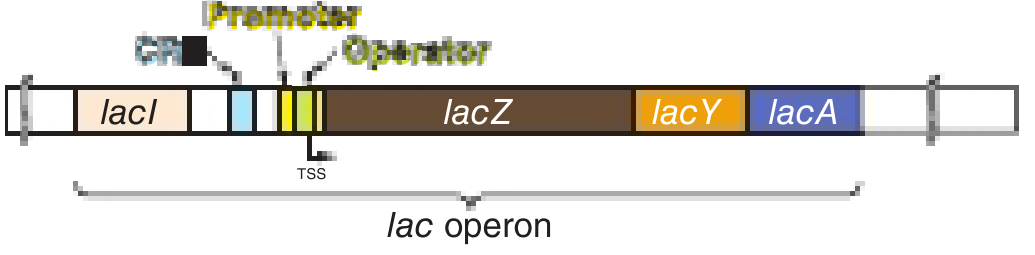
From Harper's Illustrated Biochemistry, 32nd Ed., Figure 38-2
The operon has two classes of elements:
Regulatory elements
| Element | Function |
|---|---|
| lacI gene | Constitutively transcribed; encodes the LacI repressor protein |
| CRE (cAMP Response Element) | Binding site for CAP-cAMP complex (positive regulator) |
| Promoter | RNA polymerase binding site |
| Operator | LacI repressor binding site; overlaps the promoter; a 21-bp inverted palindrome |
Structural genes
| Gene | Protein | Function |
|---|---|---|
| lacZ | β-galactosidase | Cleaves lactose into glucose + galactose; also converts lactose → allolactose |
| lacY | Galactoside permease | Transports lactose into the cell |
| lacA | Thiogalactoside transacetylase | Acetylates β-galactosides (function less clear) |
All three genes are transcribed into a single polycistronic mRNA with independent AUG/UAA codons, so each protein is translated separately (not cleaved from a precursor).
2. Structural Gene Products
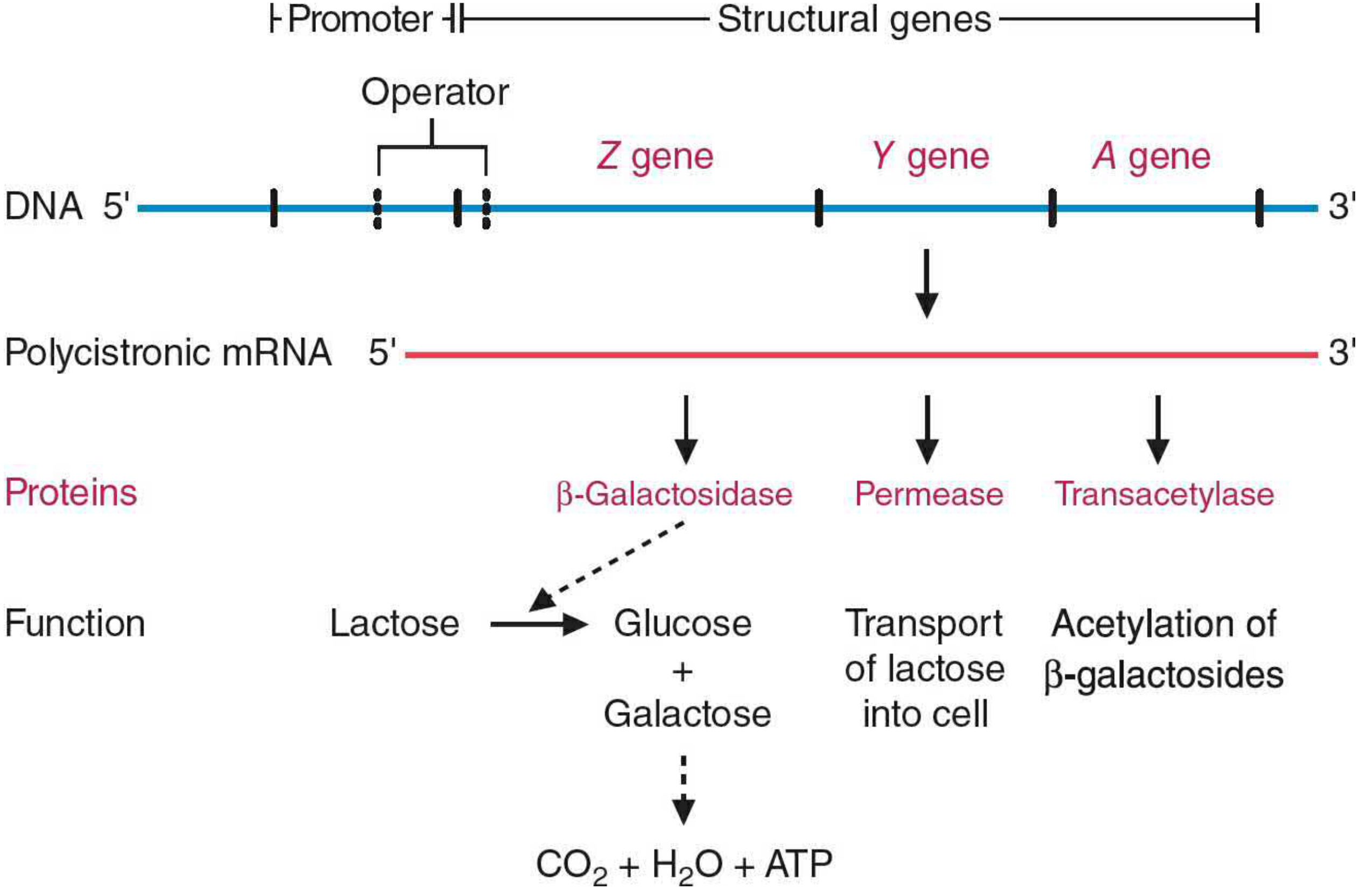
From Basic Medical Biochemistry, 6th Ed., Figure 15.5
3. Regulatory Mechanisms
The lac operon is under dual control: negative regulation by the LacI repressor and positive regulation by CAP-cAMP.
A. Negative Regulation - The LacI Repressor
- lacI is expressed constitutively, producing ~30 tetrameric LacI repressor molecules per cell
- Each tetramer has four identical subunits (MW 38,000 each)
- LacI binds the operator with very high affinity (Kd ~10⁻¹³ mol/L), occupying it >95% of the time
- When bound, LacI blocks RNA polymerase from binding the promoter → no transcription of lacZ, lacY, lacA
Key detail on the operator: It contains a 21-bp inverted palindrome:
5'-AATTGT GAG C GATAACAATT
3'-TTAACACTCG C CTATTGTTAA
Two of the four repressor subunits contact the operator, making contacts in the major groove.
B. Induction by Allolactose
When lactose is present in the environment:
- A small amount of lactose enters the cell via the low basal level of permease
- β-galactosidase converts lactose → allolactose (the true inducer)
- Allolactose binds LacI repressor → conformational change → affinity for operator drops ~10,000-fold (Kd ~10⁻⁹ mol/L)
- LacI dissociates from operator → RNA polymerase binds promoter → transcription begins
- Induction is rapid: lac mRNAs are maximally induced within ~5 min; β-galactosidase peaks within ~10 min
- Under full induction: up to 5,000 β-galactosidase molecules per cell (~1000× basal level)
IPTG (isopropyl-thiogalactoside) is a classic "gratuitous inducer" - it induces the operon but is not hydrolyzed by β-galactosidase. Widely used in molecular biology for recombinant protein expression.
4. The Three States of the Lac Operon
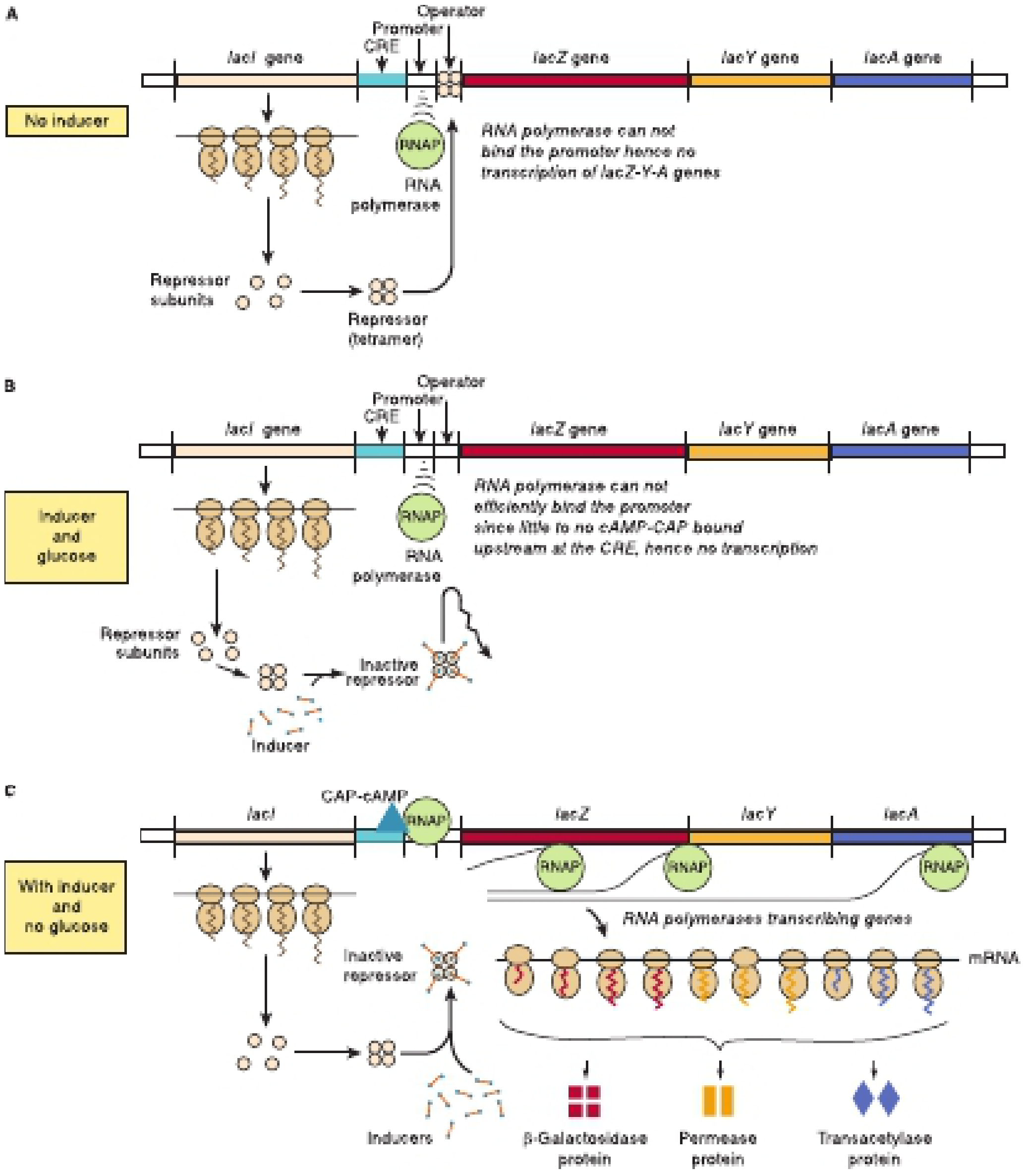
From Harper's Illustrated Biochemistry, 32nd Ed., Figure 38-3
| Condition | Repressor state | CAP-cAMP at CRE | Transcription |
|---|---|---|---|
| No lactose, glucose present | Active (bound to operator) | Absent (glucose → low cAMP) | OFF |
| Lactose + glucose | Inactive (allolactose bound) | Absent (glucose → low cAMP) | Very low |
| Lactose, no glucose | Inactive (allolactose bound) | Present (high cAMP → CAP active) | Fully ON |
5. Positive Regulation - CAP and Catabolite Repression
When glucose is present, cells preferentially use it and suppress the lac operon:
- High glucose → low cAMP levels
- CAP (Catabolite Activator Protein, also called CRP) requires cAMP to bind the CRE site
- Without CAP-cAMP at the CRE, RNA polymerase binds the promoter inefficiently → low transcription
When glucose is absent:
- cAMP levels rise
- cAMP binds CAP → conformational change → CAP-cAMP complex binds CRE upstream of promoter
- This greatly stimulates RNA polymerase binding → high-level transcription
This phenomenon (suppression of other carbon-source operons in the presence of glucose) was originally called catabolite repression, but is actually a form of positive regulation.
6. Lac Operon Expression Summary
Glucose HIGH + Lactose absent → Repressor ON, CAP OFF → No expression
Glucose HIGH + Lactose present → Repressor OFF, CAP OFF → Minimal expression
Glucose LOW + Lactose absent → Repressor ON, CAP ON → No expression
Glucose LOW + Lactose present → Repressor OFF, CAP ON → MAXIMAL expression
7. Key Mutations and Their Effects
| Mutation | Effect |
|---|---|
| lacI⁻ (no repressor) | Constitutive expression - genes always ON |
| lacIˢ (super-repressor) | Repressor can't bind allolactose; genes always OFF |
| Operator mutation (O⁻ˢ) | Repressor can't bind operator; constitutive expression |
| cya⁻ (no adenylyl cyclase, no cAMP) | Reduced expression even with inducer; CAP can't be activated |
| crp⁻ (no CAP) | Reduced expression; positive regulation lost |
8. Clinical/Biochemical Significance
- The lac operon is the founding model for understanding gene regulation, operons, and transcription factor-DNA interactions
- IPTG-inducible expression systems derived from this operon are used universally in recombinant protein production
- Allolactose acting as inducer (not lactose itself) is a key concept: the cell must first detect and metabolize the sugar before amplifying its own import machinery
Sources: Harper's Illustrated Biochemistry, 32nd Ed., pp. 433-436; Basic Medical Biochemistry: A Clinical Approach, 6th Ed., pp. 507-509; Jawetz, Melnick & Adelberg's Medical Microbiology, 28th Ed.
This is a shared conversation. Sign in to Orris to start your own chat.