resting membrane potential
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Here is a thorough explanation of the resting membrane potential, drawn from multiple physiology textbooks.
Resting Membrane Potential (RMP)
Definition
The resting membrane potential is the electrical potential difference that exists across the membrane of excitable cells (neurons, muscle) when they are not transmitting signals - i.e., between action potentials. By convention, the intracellular potential is referenced to the extracellular potential.
- Neurons: approximately -65 to -70 mV (inside negative)
- Skeletal muscle: approximately -90 mV
- Cardiac muscle: approximately -85 to -95 mV
- Erythrocytes: as low as -10 mV
The inside of the membrane is electrically negative relative to the outside.
Ion Concentrations Across the Membrane
The Na+/K+ ATPase pump establishes and maintains the following gradients (typical nerve fiber values):
| Ion | Extracellular | Intracellular |
|---|---|---|
| Na+ | 142 mEq/L | 14 mEq/L |
| K+ | 4 mEq/L | 140 mEq/L |
- Na+ is ~10x more concentrated outside
- K+ is ~35x more concentrated inside
How the RMP Is Generated - Three Contributions
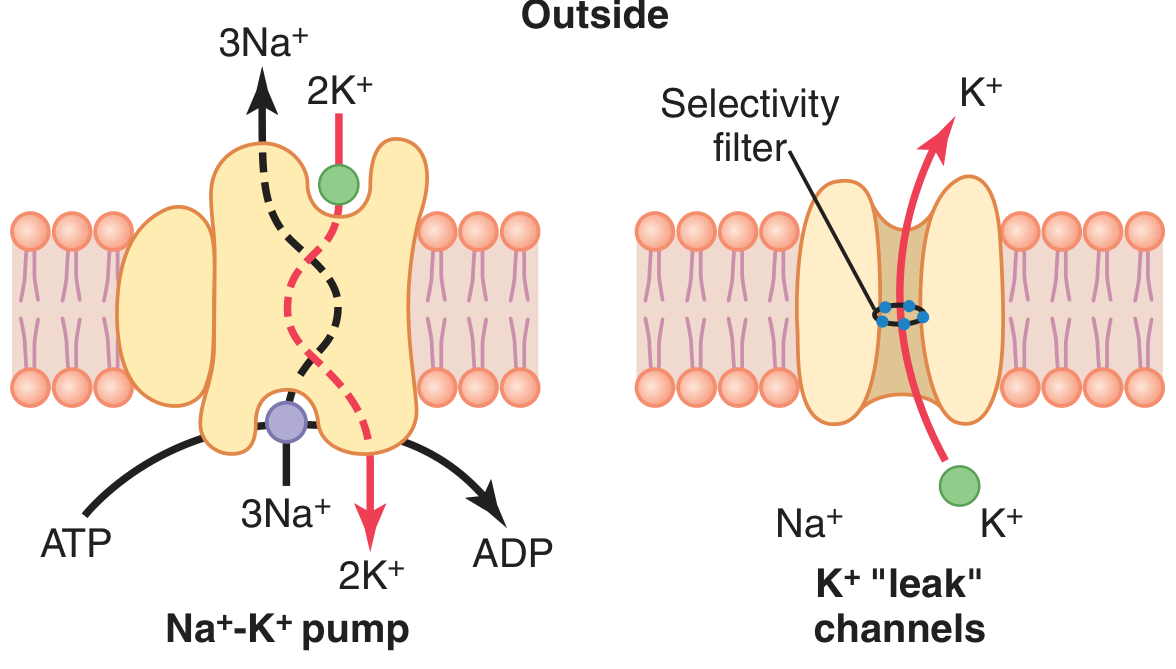
The Na+-K+ pump (left) and K+ "leak" channels (right). The selectivity filter allows K+ to exit the cell down its concentration gradient. - Guyton & Hall Textbook of Medical Physiology
1. Potassium Diffusion Potential (dominant factor)
At rest, the membrane is ~100x more permeable to K+ than to Na+, due to open K+ "leak" channels (tandem pore domain channels). K+ diffuses out of the cell down its steep concentration gradient (140 vs. 4 mEq/L). As K+ leaves, it takes positive charge with it, leaving behind large intracellular anions (proteins) that cannot cross the membrane. This creates a negative charge inside.
- The Nernst potential for K+ = -94 mV (if K+ were the only ion)
2. Small Sodium Permeability (partially offsets K+ effect)
Na+ has a slight permeability through the same K+-Na+ leak channels, diffusing inward down its gradient (142 vs. 14 mEq/L). Na+'s Nernst potential = +61 mV. Because K+ permeability so greatly exceeds Na+ permeability, the Goldman equation gives a combined potential of -86 mV.
3. The Na+/K+ ATPase Pump (small direct electrogenic contribution)
The pump transports 3 Na+ out for every 2 K+ in, creating a net loss of positive charge from the cell interior. This electrogenicity adds an additional -4 mV, bringing the total to approximately -90 mV in a nerve fiber.
The pump's more important role is indirect: it maintains the concentration gradients for K+ and Na+ that drive the diffusion potentials above. Without it, the gradients would dissipate.
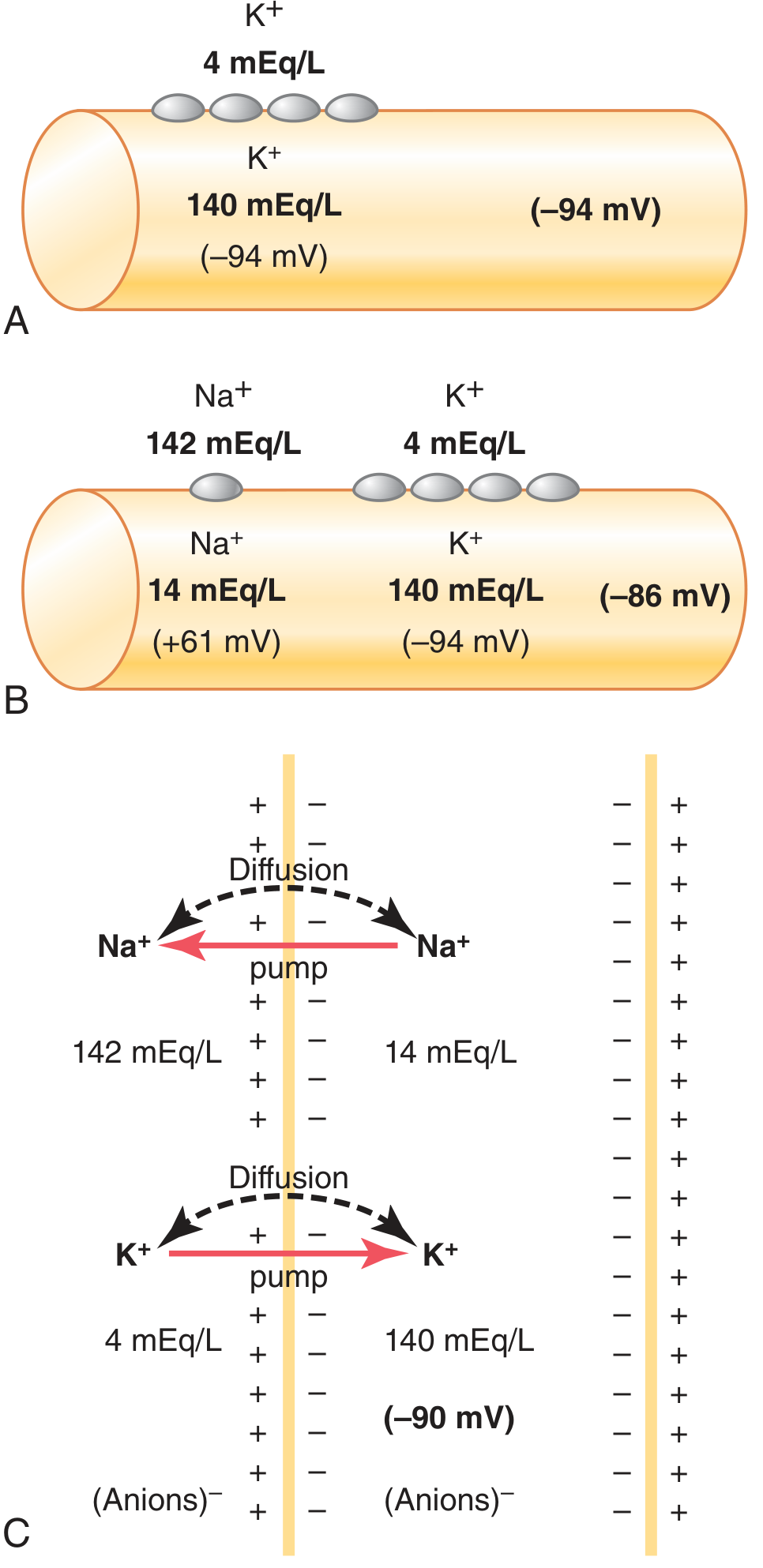
Panel A: K+ diffusion alone = -94 mV. Panel B: K+ + Na+ diffusion = -86 mV. Panel C: Adding the electrogenic Na+/K+ pump gives ~-90 mV. - Guyton & Hall, Fig. 5.5
The Nernst Equation
The equilibrium potential for any single ion is given by the Nernst equation:
E_ion = (61 mV / z) × log([ion]_outside / [ion]_inside)
Where z = valence of the ion. This is the potential at which there is no net movement of that ion - the electrical gradient exactly balances the chemical (concentration) gradient.
The Goldman Equation
When multiple ions are permeable simultaneously, the Goldman (Goldman-Hodgkin-Katz) equation calculates the membrane potential by weighing each ion's Nernst potential by its relative permeability:
V_m = (61 mV) × log [ (P_K[K+]_out + P_Na[Na+]_out + P_Cl[Cl-]_in) / (P_K[K+]_in + P_Na[Na+]_in + P_Cl[Cl-]_out) ]
At rest, P_K >> P_Na, so the RMP is dominated by K+.
Alternatively, the chord conductance equation weights each ion's equilibrium potential by its relative conductance:
E_m = (G_K / G_T) × E_K + (G_Na / G_T) × E_Na + (G_Cl / G_T) × E_Cl + ...
Equilibrium Potentials (Summary)
| Ion | Equilibrium Potential |
|---|---|
| K+ | ~-94 mV |
| Na+ | ~+61 mV |
| Cl- | ~-70 mV |
| Ca2+ | ~+132 mV |
The RMP of -70 mV sits close to E_K and E_Cl because those ions are most permeable at rest. It is far from E_Na and E_Ca2+.
Role of the Na+/K+ ATPase - Two Roles
- Direct electrogenic effect: pumps 3 Na+ out for every 2 K+ in, contributing ~-4 mV of negativity directly
- Indirect (dominant) role: maintains the K+ concentration gradient that produces the large K+ diffusion potential, and maintains the Na+ gradient that will later power the action potential upstroke
Equilibrium vs. Resting Potential
A cell at true electrochemical equilibrium for K+ would sit at -94 mV. The actual RMP is less negative (-70 to -90 mV) because:
- Small inward Na+ leak constantly depolarizes the cell
- The pump must continuously work to oppose this Na+ leak and restore gradients
This makes the resting state a steady state, not a true equilibrium.
Clinical Relevance
- Hyperkalemia (high extracellular K+): reduces the K+ concentration gradient, shifts RMP toward 0 (depolarization). This can cause muscle weakness and fatal cardiac arrhythmias.
- Hypokalemia (low extracellular K+): increases the gradient, hyperpolarizes the cell, making it harder to fire action potentials - also causing muscle weakness and arrhythmias.
- Local anesthetics (e.g., lidocaine): block Na+ channels, preventing depolarization from the RMP to threshold.
Sources:
- Costanzo Physiology, 7th Ed., pp. 25-27
- Guyton & Hall Textbook of Medical Physiology, pp. 78-79
- Neuroscience: Exploring the Brain, 5th Ed., pp. 282-287
replication of virus
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
Reading File
Reading File
Reading File
Loading Image
Loading Image
Here is a complete, textbook-grounded breakdown of viral replication.
Viral Replication
Overview
The viral replication cycle (also called the viral life cycle or lytic cycle) consists of six discrete phases:
- Attachment (Adsorption)
- Penetration (Entry)
- Uncoating
- Synthetic phase (Transcription, Translation, Genome Replication)
- Assembly (Maturation)
- Release
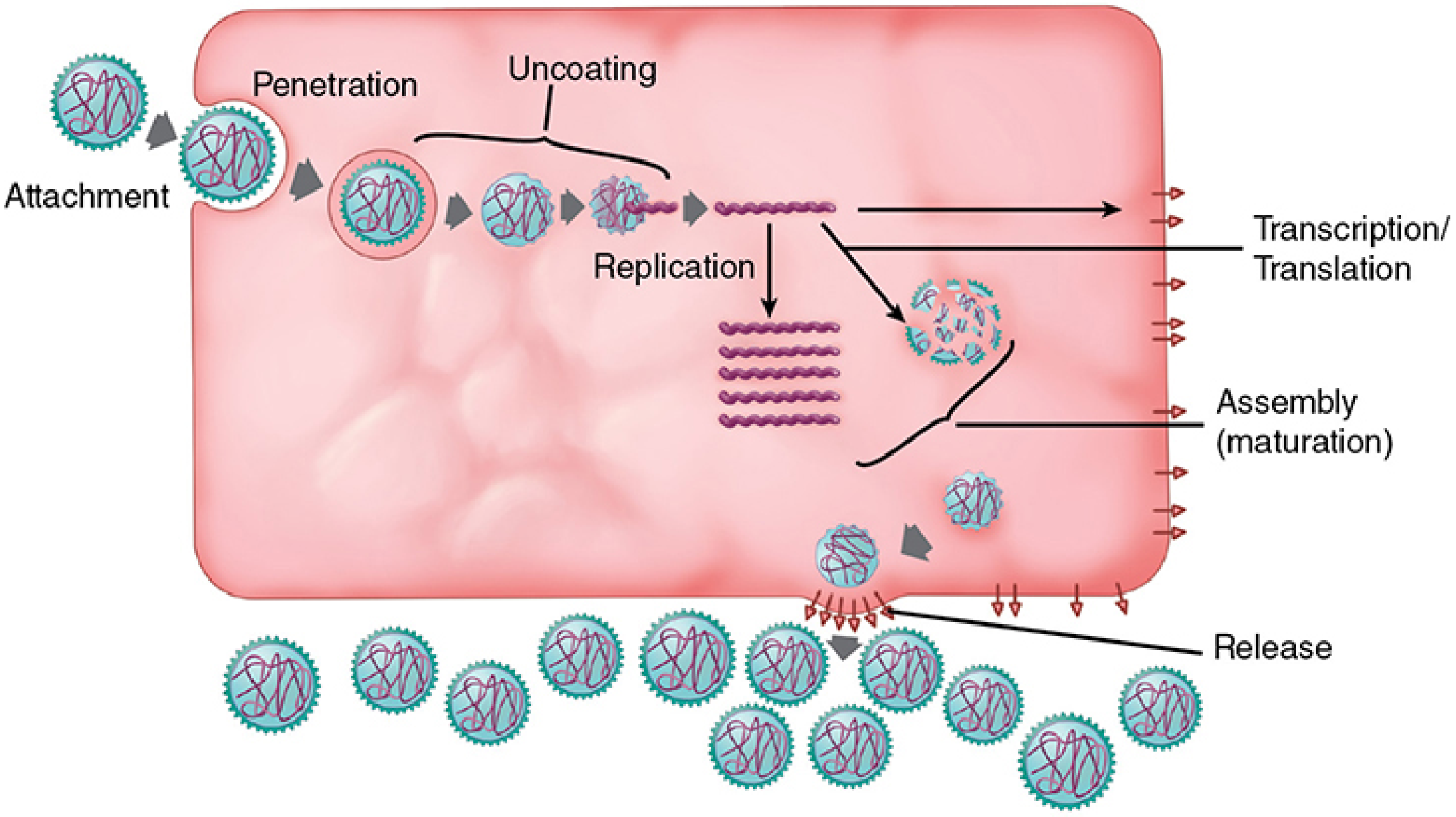
The six steps of the virus replication cycle. - Sherris & Ryan's Medical Microbiology, 8th Ed.
Step 1 - Attachment (Adsorption)
The first step is the highly specific binding of the virion to the host cell surface. Viruses have no locomotion - contact is a random diffusion event. Successful attachment depends on:
- Virion attachment proteins (VAPs) or spikes - proteins on the viral surface
- Receptors - specific molecules (usually glycoproteins) on the host cell surface; typically 10,000-100,000 per cell
This receptor specificity determines tissue tropism - which cells a virus can infect. Viruses have evolved to hijack normal cellular proteins (signaling receptors, immune components) as their receptors.
Examples of virus-receptor pairs:
| Virus | Receptor | Normal Cellular Function |
|---|---|---|
| SARS-CoV-2 (COVID-19) | ACE-2 | Angiotensin-converting enzyme 2 |
| HIV | CD4 + CCR5/CXCR4 | T-helper cell marker + chemokine receptor |
| Epstein-Barr virus | CR2 (CD21) | Complement receptor |
| Adenoviruses | Integrins | Extracellular matrix interaction |
| Influenza | Sialic acid | Glycoprotein |
Step 2 - Penetration (Entry)
After attachment, the virus enters the host cell by one of three mechanisms:
- Endocytosis - the cell engulfs the virion in a membrane-bound vesicle (endosome). Most common route.
- Membrane fusion - enveloped viruses fuse their lipid envelope directly with the plasma membrane, releasing the nucleocapsid into the cytoplasm (e.g., HIV, herpesviruses).
- Direct injection - bacteriophages inject their nucleic acid through the bacterial cell wall while the capsid remains outside.
Step 3 - Uncoating
After entry, the viral capsid is removed to expose the genome. This may occur:
- In the endosome (e.g., endosomal acidification triggers uncoating)
- In the cytoplasm
- At the nuclear pore (larger DNA viruses uncoat before entering the nucleus)
- Inside the nucleus (small DNA viruses like parvoviruses and papovaviruses enter the nucleus intact through nuclear pores, then uncoat)
Reoviruses are unusual - their contents are transferred to a lysosome where proteases partially digest the capsid and activate virion-associated enzymes needed for transcription.
Step 4 - Synthetic Phase
This is the most critical step - the virus commandeers host machinery to produce mRNAs, proteins, and new genomes.
Where does replication occur?
| Virus Type | Location |
|---|---|
| Most RNA viruses | Cytoplasm |
| Influenza, Retroviruses | Nucleus (exception among RNA viruses) |
| Most DNA viruses | Nucleus |
| Poxviruses | Cytoplasm (exception among DNA viruses - they encode their own RNA and DNA polymerases) |
4a. Transcription - Genome to mRNA
Every virus must produce mRNAs readable by host ribosomes. The strategy differs by genome type:
DNA Viruses
- Use the host's RNA polymerase II in the nucleus to transcribe viral DNA into mRNA
- Poxviruses are the exception - they carry their own RNA polymerase and replicate entirely in the cytoplasm
- Larger DNA viruses (herpesviruses, adenoviruses) also encode their own DNA polymerase - this is the basis of antiviral drugs like acyclovir (targets herpesvirus thymidine kinase + DNA polymerase)
(+) Positive-Sense RNA Viruses (e.g., Picornaviruses, Coronaviruses, Flaviviruses, Togaviruses)
- Genome directly acts as mRNA - translated immediately on entry
- One of the first proteins translated is RNA-dependent RNA polymerase (RdRp), which then:
- Makes a (-) strand intermediate
- Uses it as template to synthesize more (+) strand mRNAs and new genomes
(-) Negative-Sense RNA Viruses (e.g., Rhabdoviruses, Paramyxoviruses, Orthomyxoviruses, Filoviruses)
- Genome is antisense to mRNA and cannot be directly translated
- Must carry a virion-associated RNA-dependent RNA polymerase (transcriptase) packaged inside the virion
- This transcriptase immediately transcribes the (-) genome into (+) mRNAs upon entry
Retroviruses (e.g., HIV) - Special Case
- (+) sense RNA genome, but is not translated directly
- Virion carries reverse transcriptase (RNA-dependent DNA polymerase)
- Steps:
- Reverse transcriptase copies RNA genome → (-) DNA strand
- RNase H activity of reverse transcriptase degrades the RNA template
- (+) DNA strand synthesized → double-stranded DNA (dsDNA)
- dsDNA integrates into the host chromosome (provirus) via integrase
- Host RNA polymerase transcribes the integrated provirus → viral mRNAs + new genomic RNA
Double-Stranded RNA Viruses (e.g., Reoviruses)
- Must also carry a virion-associated transcriptase (host cells have no dsRNA polymerase)
4b. Translation
- Viral mRNAs are translated by host ribosomes
- Human cells require monocistronic mRNA (one protein per mRNA), unlike bacteria
- Viruses solve this by: polyprotein precursors cleaved by viral proteases (e.g., picornaviruses, HIV), subgenomic RNAs, or ribosomal frameshifting
Temporal regulation of gene expression:
- Early proteins: enzymes needed for genome replication (polymerases, kinases)
- Late proteins: structural proteins (capsid, envelope) needed for assembly
4c. Genome Replication
- New genomic copies are made using the appropriate polymerase
- RNA viruses encode their own RNA-dependent RNA polymerase (replicase) for this
- The same template is used for both transcription (mRNAs) and replication (full-length genomes) - these processes must be temporally separated
- DNA virus genome replication mirrors host DNA replication, but with varying degrees of dependence on host machinery:
- Parvoviruses (smallest DNA viruses): fully dependent on host - require dividing cells (S-phase)
- Polyomaviruses/Papillomaviruses: encode origin-binding protein, rest done by host
- Herpesviruses/Adenoviruses: encode their own DNA polymerase + accessory proteins
- Poxviruses (largest): encode all replication machinery themselves
Step 5 - Assembly (Maturation)
New viral genomes are packaged with structural proteins to form progeny virions.
Helical capsids (e.g., TMV, influenza): Doughnut-shaped protein disks are added stepwise to the RNA genome, elongating in both directions from a specific packaging site. The process ceases when the RNA ends are reached.
Icosahedral capsids (e.g., adenoviruses, picornaviruses): The hollow capsid is preassembled first (by aggregation of pentamers and hexamers), then the genome is threaded in - or a small protein-genome complex nucleates capsid assembly around itself.
Enveloped viruses: The nucleocapsid must also acquire a lipid envelope - this happens during release (see below).
Step 6 - Release
Naked (non-enveloped) viruses
- Released by cell lysis (cell death)
- Cell death results from viral takeover of cellular machinery, triggering apoptosis
- Some viruses (e.g., picornaviruses) shut off host protein synthesis early; DNA viruses interfere with cell-cycle controls
- Some viruses encode proteins that delay apoptosis to allow more time for replication
Enveloped viruses - Budding
- Released by budding from a cellular membrane - coupled with final envelope acquisition
The budding sequence is:
- Viral glycoprotein spikes are synthesized and inserted into the plasma membrane (via Golgi pathway)
- The matrix (M) protein coats the cytoplasmic side of the membrane at the spike cluster
- The assembled nucleocapsid is attracted to the matrix protein
- The membrane wraps around the nucleocapsid and pinches off, releasing the complete virion
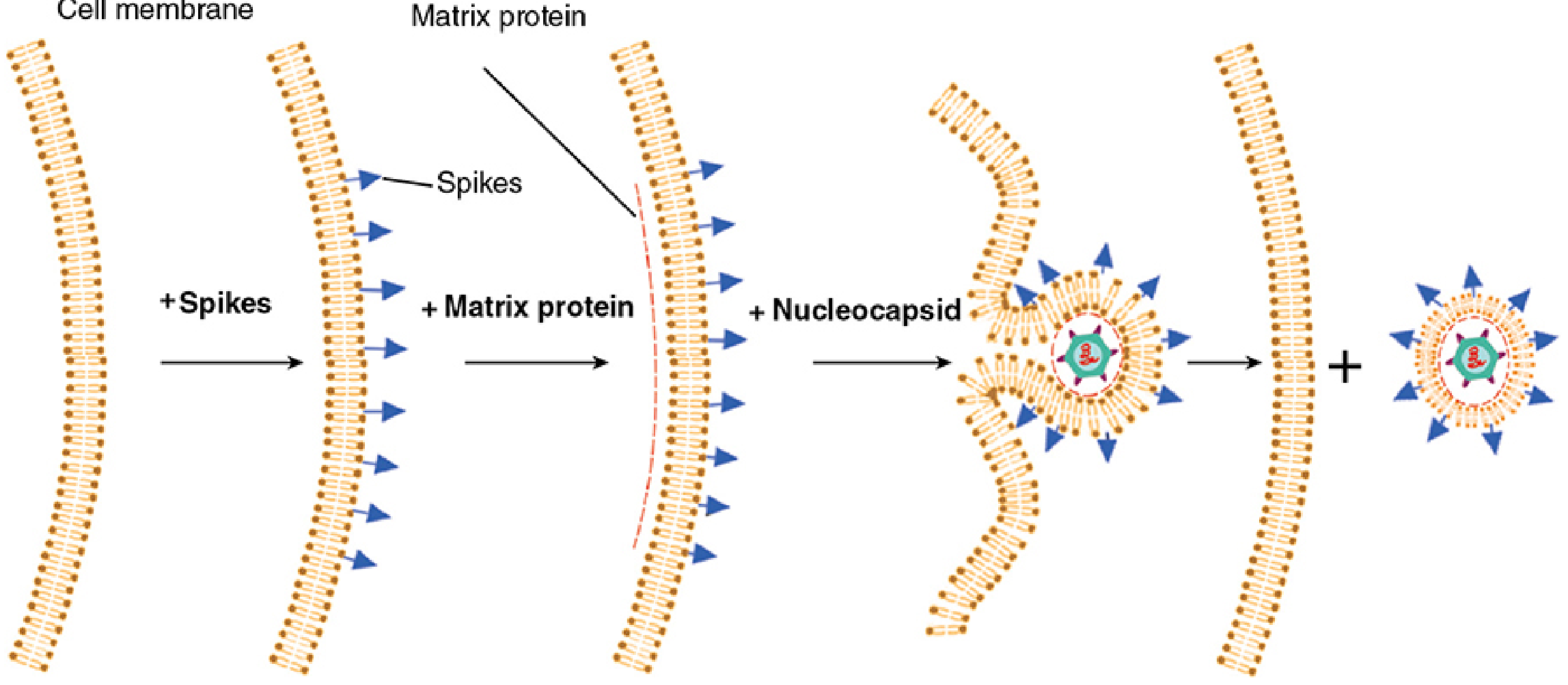
Viral release by budding: spikes insert into the plasma membrane → matrix protein accumulates → nucleocapsid is recruited → membrane wraps and buds off. - Sherris & Ryan's Medical Microbiology, 8th Ed.
Where budding occurs:
- Most enveloped viruses: plasma membrane
- Herpesviruses: nuclear membrane (then exit via exocytic vesicles)
- Coronaviruses, poxviruses: cytoplasmic membranes (Golgi apparatus)
Initial budding usually doesn't kill the cell, but the accumulation of many daughter viruses eventually disrupts membrane integrity. HIV is an exception - it is cytopathic even as a retrovirus, killing CD4+ T cells through multiple mechanisms.
Outcomes of Infection
| Outcome | Description | Example |
|---|---|---|
| Productive (lytic) | Virus replicates, cell dies, progeny released | Poliovirus, influenza |
| Persistent/Chronic | Low-level replication, little cell damage | HCV, HBV |
| Latent | Viral DNA maintained silently, no replication | HSV, VZV, EBV |
| Abortive | Cell doesn't support replication; early proteins may cause cell death | Some adenovirus infections |
| Oncogenic transformation | Nonproductive; viral DNA integrates, drives cell proliferation | HPV, EBV |
Clinical Relevance - Antiviral Drug Targets
Understanding each replication step reveals specific drug targets:
| Step | Target | Example Drug |
|---|---|---|
| Attachment | Viral spikes or host receptor | Maraviroc (CCR5 blocker - HIV) |
| Fusion | Viral fusion protein | Enfuvirtide (HIV) |
| Reverse transcription | Reverse transcriptase | AZT, Tenofovir (HIV) |
| Integration | Integrase | Raltegravir (HIV) |
| DNA replication | Viral DNA polymerase | Acyclovir (HSV) |
| Protease cleavage | Viral protease | Lopinavir (HIV) |
| Release | Neuraminidase (influenza) | Oseltamivir (Tamiflu) |
Sources:
- Sherris & Ryan's Medical Microbiology, 8th Ed., Ch. 6 (pp. 198-231)
- Jawetz, Melnick & Adelberg's Medical Microbiology, 28th Ed.
how does uncoating occur
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
Excellent content retrieved. Here is the complete explanation.
Viral Uncoating
What Is Uncoating?
Uncoating is the process by which a virus sheds its protein coat (capsid) and/or envelope after entering the host cell, releasing the viral genome so it can be transcribed and replicated. It marks the start of the eclipse period - the interval during which no intact infectious virus can be detected, because the virion has been dismantled but new progeny have not yet been assembled.
The mechanism of uncoating varies significantly depending on whether the virus is:
- Enveloped or naked (non-enveloped)
- Entering by direct fusion or by endocytosis
- DNA or RNA, and where replication occurs
Mechanism 1 - Direct Fusion at the Plasma Membrane (Enveloped Viruses)
Some enveloped viruses bypass the endosome entirely. Their surface fusion proteins interact with host receptors at neutral pH at the plasma membrane and trigger direct fusion of the viral envelope with the cell membrane. The nucleocapsid is released directly into the cytoplasm - this is uncoating in a single step.
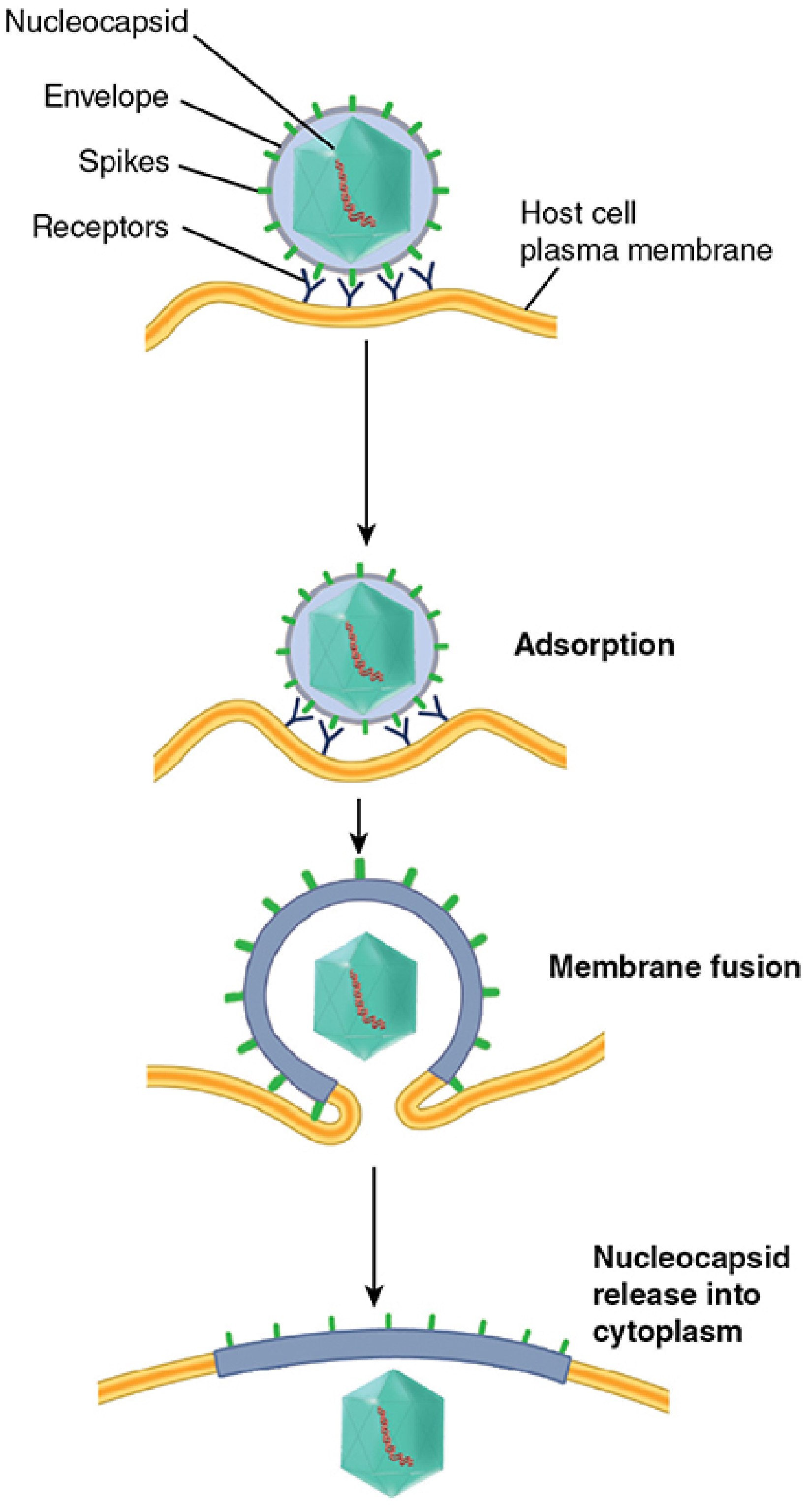
Adsorption → direct membrane fusion → nucleocapsid released into cytoplasm. - Sherris & Ryan's Medical Microbiology, 8th Ed.
Examples:
- Paramyxoviruses (measles, RSV, mumps): the F1 (fusion) protein mediates fusion at neutral pH at the plasma membrane. If the F0 precursor is not cleaved to F1, no fusion occurs and infection fails.
- HIV and some retroviruses: gp41 (fusion protein) drives fusion with the plasma membrane after gp120 binds CD4 + co-receptor (CCR5/CXCR4).
- Herpesviruses: fuse at the plasma membrane, releasing a tegument-coated capsid into the cytoplasm.
A side effect of this: because the viral envelope merges with the plasma membrane, infected cells display viral glycoproteins on their surface and tend to fuse with neighboring uninfected cells - producing syncytia (multinucleated giant cells). This is a hallmark of paramyxovirus and HIV infections.
Mechanism 2 - Viropexis (Receptor-Mediated Endocytosis) + Low pH Uncoating
This is the most common mechanism, used by both enveloped and naked capsid viruses. The process is called viropexis.
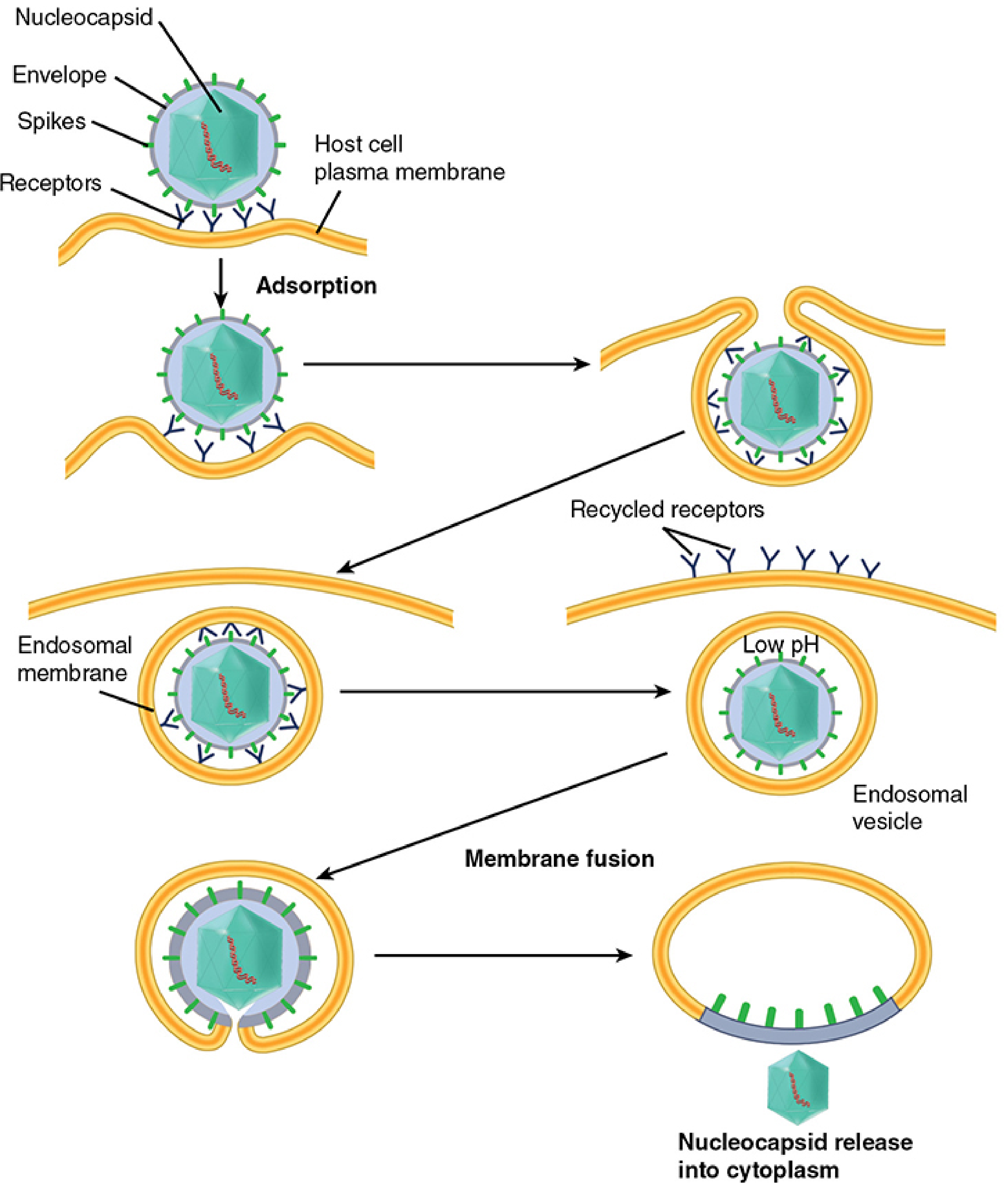
Left path: direct fusion. Right path (viropexis): adsorption → endosomal vesicle formation → receptors recycled → endosome acidified (low pH) → membrane fusion → nucleocapsid released into cytoplasm. - Sherris & Ryan's Medical Microbiology, 8th Ed.
Steps:
- Virus adsorbs to the cell surface via receptor binding
- The plasma membrane invaginates and engulfs the virion - enclosing it in a cytoplasmic endosomal vesicle
- Cell surface receptors are recycled back to the membrane
- The endosome is acidified by cellular vacuolar H+-ATPases (pH drops to ~5-6)
- Low pH triggers a conformational change in a viral spike protein
- This drives fusion of the viral envelope with the endosomal membrane, releasing the nucleocapsid into the cytoplasm
Examples using this route:
- Influenza virus: the hemagglutinin (HA) undergoes acid-induced conformational change in the endosome, driving fusion. Simultaneously, the M2 ion channel allows H+ to flow into the virion interior, disrupting matrix protein-nucleocapsid interactions and completing uncoating. This is the target of amantadine and rimantadine (M2 blockers) - blocking H+ influx through M2 prevents the matrix proteins from dissociating, so uncoating fails.
- Togaviruses (rubella), rhabdoviruses (rabies), coronaviruses: endocytic entry with pH-triggered fusion.
Uncoating of Naked Capsid Viruses via Viropexis
Naked viruses cannot escape the endosome by membrane fusion (they have no envelope). Instead:
- Poliovirus and picornaviruses: low pH in the endosome causes capsid proteins to expose hydrophobic domains. These domains bind to the endosomal membrane, creating a pore through which the RNA genome is injected directly into the cytoplasm. Drug pleconaril and related compounds (arildone, disoxaril) bind to a hydrophobic cleft in the receptor-binding "canyon" on the picornavirus capsid and block this disassembly step.
- Adenovirus: taken up by endocytosis; endosomal acidification (half-life ~5 minutes) triggers escape of ~90% of particles into the cytosol. Uncoating is sequential and organized - it systematically breaks the stabilizing interactions built during viral maturation. Microtubules transport the partially uncoated particle to the nucleus, where final uncoating and DNA release occur, possibly at the nuclear membrane.
- Reovirus (unusual): the endosome contents are transferred to a lysosome, where lysosomal proteases strip away outer capsid proteins and activate virion-associated enzymes needed for transcription. The inner core particle (not the naked genome) is then released.
Mechanism 3 - Bacteriophage Injection (No Endocytosis)
Bacteriophages use a completely different strategy: penetration and uncoating are simultaneous, and the capsid never enters the cell.
- Tailed phages (e.g., T4) attach via tail fibers to the bacterial cell wall
- The hollow tail acts as a syringe, injecting only the nucleic acid genome into the cell
- The protein capsid remains outside, shed at the surface
- A small number of virion proteins tightly associated with the DNA may enter alongside the genome
Mechanism 4 - Two-Stage Uncoating in Poxviruses
Poxviruses have the most complex uncoating process because they are the largest DNA viruses and replicate entirely in the cytoplasm:
Stage 1 - Primary uncoating:
- The virion envelope fuses with the plasma membrane (or endosomal membrane), releasing the viral core into the cytoplasm
- The core remains intact with its own internal enzymes
Stage 2 - Secondary uncoating (requires viral gene expression):
- The viral RNA polymerase (carried inside the core) transcribes ~half the genome into early mRNAs, which exit the core into the cytoplasm
- Among the early proteins translated is an "uncoating protein"
- This protein acts on the cores to complete uncoating and liberate the viral DNA from the core
- This step requires both RNA and protein synthesis - making it unique among viruses
Herpesvirus Uncoating - Nuclear Targeting
Herpesviruses enter by direct fusion but their uncoating is staged:
- Envelope fuses with plasma membrane → tegument-coated capsid released into cytoplasm
- Tegument proteins are shed
- The capsid travels along microtubules to the nucleus
- At the nuclear pore, the capsid docks and the DNA is injected into the nucleus
- The viral DNA immediately circularizes upon release from the capsid
- The empty capsid (ghost) remains at the nuclear pore
Summary Table
| Virus | Entry Route | Trigger for Uncoating | Where Genome is Released |
|---|---|---|---|
| Paramyxoviruses (measles, RSV) | Direct fusion at plasma membrane | Neutral pH - F protein conformational change | Cytoplasm |
| HIV | Direct fusion at plasma membrane | Receptor binding + co-receptor | Cytoplasm, then nucleus |
| Influenza | Endocytosis (viropexis) | Endosomal low pH + M2 H+ channel | Cytoplasm |
| Poliovirus | Endocytosis (viropexis) | Low pH - hydrophobic domain exposure | Cytoplasm via membrane pore |
| Adenovirus | Endocytosis (viropexis) | Endosomal acidification | Cytoplasm + nuclear pore |
| Reovirus | Endocytosis → lysosome | Lysosomal proteases | Cytoplasm (as core particle) |
| Herpesvirus | Direct fusion | Nuclear pore docking | Nucleus |
| Poxvirus | Fusion + 2-stage process | Viral "uncoating protein" (requires protein synthesis) | Cytoplasm |
| Bacteriophages | Direct injection | None - capsid stays outside | Bacterial cytoplasm |
Clinical Relevance - Drugs That Block Uncoating
| Drug | Virus | Mechanism |
|---|---|---|
| Amantadine / Rimantadine | Influenza A | Block the M2 H+ channel → matrix proteins don't dissociate from nucleocapsid → uncoating fails |
| Pleconaril / Arildone | Picornaviruses (enteroviruses, rhinoviruses) | Bind the hydrophobic canyon in the capsid → prevent conformational change needed for genome release |
| Enfuvirtide (T20) | HIV | Inhibits gp41 fusion peptide → blocks envelope-membrane fusion = blocks entry/uncoating |
| Docosanol | Herpes simplex | Inhibits fusion of enveloped viruses with cellular membranes |
Sources:
- Sherris & Ryan's Medical Microbiology, 8th Ed., pp. 208-213
- Jawetz, Melnick & Adelberg's Medical Microbiology, 28th Ed.
- Murray's Medical Microbiology, 9th Ed., pp. 455-456
what happens at 4 5 6 step of replication explain from basic
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Reading File
Reading File
All the content I need is in hand. Here is the full explanation from the basics.
Steps 4, 5, and 6 of Viral Replication - From the Basics
Quick Recap of Steps 1-3
Before steps 4-6 make sense, remember what happened first:
- Step 1 - Attachment: virus stuck to the host cell surface
- Step 2 - Penetration: virus entered the cell
- Step 3 - Uncoating: the protein coat was removed, releasing the bare viral genome
Now the naked genome is inside the cell. Steps 4, 5, and 6 are about building new viruses from scratch using the host cell's machinery.
STEP 4 - The Synthetic Phase (Making Viral Parts)
This is the most complex step. Think of it as the virus "giving instructions" to the cell to become a factory that builds virus parts. Three things need to be made:
- mRNAs (instructions for making proteins)
- Viral proteins (structural + non-structural)
- New viral genomes (copies of the viral DNA/RNA)
4A - Transcription: Making mRNA from the Viral Genome
What is mRNA? - mRNA is a molecular message. The cell's ribosomes (protein-building machines) can only read mRNA. So the viral genome - whether DNA or RNA - must first be converted into mRNA before any viral protein can be made.
For DNA Viruses (e.g., Herpesviruses, Adenoviruses, Papillomaviruses)
- The viral DNA travels to the nucleus
- The host cell's own RNA polymerase II reads the viral DNA and transcribes it into viral mRNA
- The mRNAs travel out to the cytoplasm where ribosomes translate them into proteins
- Exception - Poxviruses: they are so large they carry their own RNA polymerase inside the virion and replicate entirely in the cytoplasm, never needing the nucleus
For Positive-Sense (+) RNA Viruses (e.g., Poliovirus, Coronavirus, Hepatitis C)
- The genome IS already mRNA - it has the same polarity (+ sense = readable directly)
- As soon as it enters the cytoplasm, ribosomes attach directly to the genome and translate it immediately - no transcription step needed first
- The first proteins made include RNA-dependent RNA polymerase (RdRp) - this is the enzyme that will later copy the RNA genome
For Negative-Sense (-) RNA Viruses (e.g., Influenza, Rabies, Ebola, Measles)
- The genome is the opposite polarity to mRNA - it cannot be read by ribosomes directly
- The virus solves this by carrying its own RNA-dependent RNA polymerase (RdRp) already packaged inside the virion
- As soon as uncoating occurs, this pre-packaged polymerase immediately begins copying the (-) strand genome into (+) strand mRNAs
- These mRNAs are then translated by host ribosomes
For Retroviruses (e.g., HIV)
- The genome is (+) RNA, but retroviruses do NOT translate it directly
- Instead, the virion carries reverse transcriptase - it copies RNA → DNA
- Step 1: RNA → single-stranded (-) DNA
- Step 2: RNA template degraded by RNase H activity
- Step 3: (+) DNA strand made → double-stranded DNA (dsDNA)
- The dsDNA enters the nucleus and is integrated into the host chromosome (provirus) by the enzyme integrase
- Now the host's RNA polymerase transcribes the integrated provirus - producing both mRNAs AND new genomic RNA
Temporal Regulation: Early vs. Late Genes
Viruses do not make all proteins at once - gene expression is timed in waves:
| Phase | What's Made | Purpose |
|---|---|---|
| Immediate early (α) | Transcription factors, regulatory proteins | Take over the cell; shut down host defenses |
| Early (β) | Enzymes - especially DNA/RNA polymerases | Replicate the viral genome |
| Late (γ) | Structural proteins (capsid, envelope glycoproteins) | Used for assembly of new virions |
This timing ensures the cell is "prepared" and genome copies are available before costly structural proteins are manufactured.
4B - Genome Replication: Making Copies of the Viral Genome
After the first wave of early proteins are made (including polymerases), the virus begins copying its genome to provide DNA/RNA for all the new daughter virions.
-
DNA viruses: DNA replication in the nucleus; uses viral DNA polymerase (or borrows host polymerase for smaller viruses). Follows the same biochemical rules as host DNA replication but requires a primer.
- Small DNA viruses (parvoviruses): fully depend on host - require cells that are actively dividing (in S phase)
- Medium DNA viruses (papillomaviruses, adenoviruses): encode origin-binding protein; use a mix of host and viral machinery
- Large DNA viruses (herpesviruses): encode their own complete DNA polymerase + accessory proteins
- Largest (poxviruses): encode everything themselves
-
RNA viruses: use the RNA-dependent RNA polymerase (RdRp) = replicase to copy RNA → RNA
- (+) strand viruses: genome copied to a (-) strand intermediate, then back to many (+) strands
- (-) strand viruses: genome copied to a full-length (+) strand (antigenome), then back to many (-) strands for packaging
-
The host cell has no RdRp - it cannot copy RNA from an RNA template. This is why all RNA viruses must bring or make their own polymerase.
4C - Translation: Making Viral Proteins
Viral mRNAs are read by host ribosomes in the cytoplasm. Two types of proteins are made:
Non-structural (functional) proteins:
- Polymerases / replicases
- Proteases (cleave polyproteins into individual proteins - e.g., HIV protease)
- Kinases, integrases
- Proteins that shut off host cell protein synthesis or immune defenses
Structural proteins:
- Capsid proteins (VP1, VP2, VP3... in picornaviruses)
- Envelope glycoproteins (inserted into ER membrane, processed through Golgi, travel to cell surface)
- Matrix (M) proteins (link envelope to nucleocapsid)
- Nucleoproteins (coat the viral RNA)
The Monocistronic Rule
Human ribosomes can only translate one protein per mRNA (monocistronic). Viruses solve this in different ways:
- DNA viruses: splice a single large precursor RNA into many separate mRNAs (using nuclear splicing machinery)
- Segmented RNA viruses (influenza, rotavirus): each genome segment = one gene = one mRNA
- (-) strand RNA viruses: polymerase starts a new mRNA at the beginning of each gene
- (+) strand RNA viruses (picornaviruses, flaviviruses): translate one giant polyprotein, then a viral protease cuts it into individual functional proteins
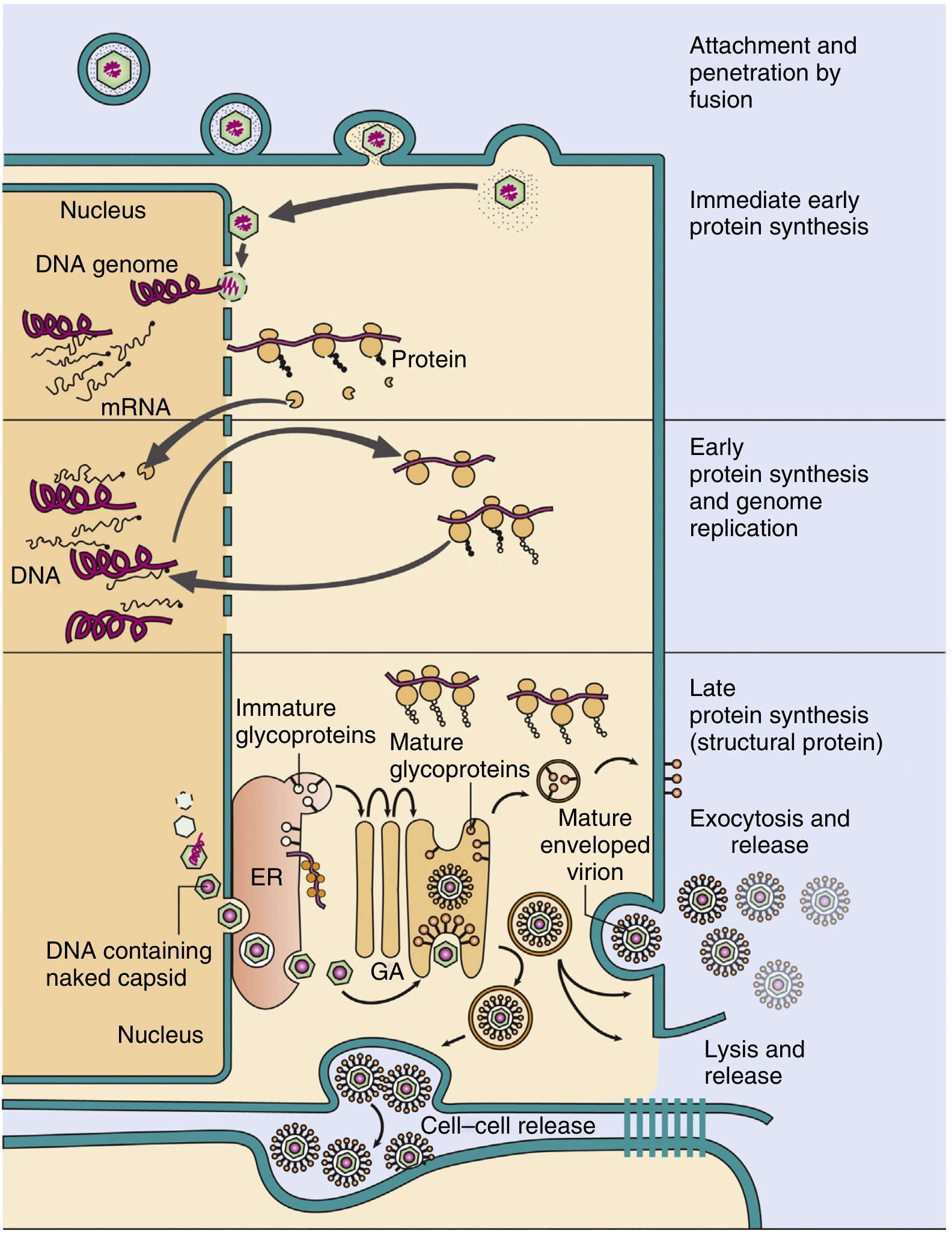
Complete replication cycle of herpes simplex virus (HSV): attachment and fusion → immediate early protein synthesis → early proteins + genome replication → late structural proteins → assembly (capsids in nucleus, then ER/Golgi for envelope) → three release routes: exocytosis, lysis, or cell-cell bridges. ER = endoplasmic reticulum; GA = Golgi apparatus. - Murray's Medical Microbiology, 9th Ed.
STEP 5 - Assembly (Packaging / Maturation)
Once enough viral parts are made, they must be put together into complete new virions. This is called assembly or encapsidation.
Four Basic Rules of Assembly
- Self-assembly - the proteins have complementary shapes and naturally come together (like puzzle pieces)
- Stepwise and ordered - cannot skip steps; each piece must attach before the next one can
- Pre-formed capsomeres - individual protein subunits first cluster into intermediate capsomere units, which then join to form the full capsid
- Packaging signal - the viral genome contains a specific sequence called a packaging site that the assembling capsid recognizes, ensuring only the correct genome gets packaged (not random cellular RNA or DNA)
Assembly of Helical Capsids (e.g., TMV, Influenza, Paramyxoviruses)
- Protein subunits are pre-formed into doughnut-shaped disks
- These disks attach to the viral RNA at a specific packaging site
- They add stepwise in both directions along the RNA like beads on a string
- The RNA and protein together coil into a helix
- The process automatically stops when the end of the RNA is reached
- For segmented helical viruses (influenza), each genome segment is assembled separately into its own nucleocapsid, then all segments are gathered together during final virion assembly
Assembly of Icosahedral (Cubic) Capsids (e.g., Adenovirus, Picornavirus, Herpesviruses)
- Individual capsid proteins first cluster into pentamers (groups of 5) and hexamers (groups of 6)
- These capsomeres condense together to form an empty hollow capsid shell
- The viral genome is then threaded into the empty shell (DNA viruses) - OR
- A small nucleation complex of capsid protein + genome forms first, then the rest of the capsid assembles around it (some RNA viruses)
Where Assembly Occurs
| Virus | Where Assembly Happens |
|---|---|
| Most DNA viruses (herpesviruses, adenoviruses) | Nucleus - capsid proteins migrate in, assemble around DNA |
| Most RNA viruses (picornaviruses, paramyxoviruses) | Cytoplasm |
| Poxviruses | Cytoplasm (in specialized "viral factories") |
Enveloped Viruses - Acquiring the Lipid Envelope
Naked (non-enveloped) viruses are complete after the nucleocapsid assembles. Enveloped viruses have an extra step - they must wrap themselves in a lipid membrane studded with viral glycoproteins.
How the envelope is acquired:
- Viral envelope glycoproteins are synthesized by ribosomes on the rough ER
- They travel through the Golgi apparatus where they are glycosylated (sugar groups added) and processed into their mature form
- They are inserted into the desired membrane (plasma membrane, ER membrane, nuclear membrane, or Golgi membrane depending on virus)
- The nucleocapsid recognizes and migrates to these membrane patches
- The membrane wraps around the nucleocapsid → budding → complete enveloped virion
For herpesviruses specifically: capsid assembles in the nucleus → buds through the nuclear membrane (acquiring an initial envelope) → travels through the ER and trans-Golgi network → acquires final tegument proteins and definitive glycoprotein envelope → released by exocytosis.
STEP 6 - Release (Exit from the Host Cell)
Once assembled, the new virions must escape from the cell to infect other cells. The mechanism depends entirely on whether the virus is enveloped or naked.
Release of Naked Capsid Viruses - Cell Lysis
Naked viruses (poliovirus, adenovirus, rhinovirus) accumulate inside the cell and are released by lysis - the cell bursts open.
How cell death occurs:
- The virus hijacks and shuts down normal cellular functions (e.g., picornaviruses block host protein synthesis)
- Viral proteins may disrupt the cell membrane or interfere with cell-cycle control
- This triggers apoptosis (programmed cell death) - the cell essentially destroys itself
- Some viruses encode anti-apoptotic proteins to delay this (to maximize virion production before the cell dies)
- Cell lysis releases hundreds to thousands of new virions into the extracellular environment at once
Each infected cell can produce up to 100,000 particles - though only 1-10% may be fully infectious (defective particles result from assembly errors).
Release of Enveloped Viruses - Budding
Most enveloped viruses are released by budding - the virus pushes out through the cell membrane taking a piece of lipid bilayer with it as its envelope. This process does NOT immediately kill the cell.
The budding sequence:
- Viral glycoprotein spikes cluster in a region of the plasma membrane (delivered there via the Golgi pathway)
- The cytoplasmic face of that membrane patch becomes coated with matrix (M) protein
- The completed nucleocapsid is attracted to and docks with the matrix protein
- The membrane curves around the nucleocapsid and pinches off, releasing a complete enveloped virion outside the cell
- The cell membrane is repaired - the cell survives and can continue budding out more virions
Where budding occurs:
- Most enveloped viruses (influenza, HIV, paramyxoviruses): plasma membrane
- Herpesviruses: nuclear membrane (then transported out via exocytosis)
- Coronaviruses, poxviruses: cytoplasmic/Golgi membranes
Important consequence of budding: Viral glycoproteins remain on the surface of the infected cell's plasma membrane. The immune system (cytotoxic T cells) recognizes these foreign proteins and attacks infected cells before they finish releasing virions - this is actually a key antiviral defense mechanism.
Special Case - HIV (Cytopathic Retrovirus)
Most retroviruses bud without killing the cell. HIV is an exception - it is cytopathic and kills CD4+ T cells through:
- Accumulation of unintegrated viral DNA in the cytoplasm (toxic)
- Toxic effects of certain viral proteins (gp120, Vpr)
- Alterations in cell membrane permeability
- Induction of apoptosis
- Cell-cell fusion (syncytia formation)
Summary Diagram
STEP 4 - SYNTHETIC PHASE
|
├── Transcription: Genome → mRNA
| • DNA viruses: host RNA pol II (in nucleus)
| • (+) RNA: genome = mRNA, translate directly
| • (-) RNA: virion-packaged RdRp makes mRNA
| • Retroviruses: reverse transcriptase RNA→DNA→integrated→mRNA
|
├── Translation: mRNA → Proteins (by host ribosomes)
| • Immediate early proteins (regulatory)
| • Early proteins (polymerases, enzymes)
| • Late proteins (capsid, glycoproteins)
|
└── Genome Replication: New copies of viral genome
• DNA viruses: DNA polymerase in nucleus
• RNA viruses: RdRp in cytoplasm
STEP 5 - ASSEMBLY
|
├── Capsid proteins self-assemble stepwise (capsomeres → capsid)
├── Genome packaged into capsid via packaging signal
├── Enveloped viruses: glycoproteins processed through Golgi →
| inserted into membrane → nucleocapsid recruits matrix protein →
| membrane wraps around nucleocapsid
└── Maturation complete: virion ready
STEP 6 - RELEASE
|
├── Naked viruses: cell LYSIS → all virions released at once
└── Enveloped viruses: BUDDING → continuous release without
immediate cell death
Sources:
- Sherris & Ryan's Medical Microbiology, 8th Ed., pp. 213-231
- Murray's Medical Microbiology, 9th Ed.
- Harrison's Principles of Internal Medicine, 22nd Ed., Ch. 19
This is a shared conversation. Sign in to Orris to start your own chat.