oxidative phosphorylation
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Reading File
Here is a comprehensive overview of oxidative phosphorylation, drawn primarily from Basic Medical Biochemistry - A Clinical Approach, 6e and Biochemistry, 8th ed (Lippincott Illustrated Reviews).
Oxidative Phosphorylation
Overview
Oxidative phosphorylation (OxPhos) is the process by which energy released from fuel oxidation is captured as ATP. Most cellular fuel oxidation (TCA cycle, fatty acid oxidation, glycolysis) produces the reduced coenzymes NADH and FAD(2H). These donate electrons to the electron transport chain (ETC), which transfers them to O2, producing H2O. The energy released is used to pump protons across the inner mitochondrial membrane, creating an electrochemical gradient that drives ATP synthesis.
The net yield is approximately:
- 2.5 mol ATP per mol NADH oxidized
- 1.5 mol ATP per mol FAD(2H) oxidized
Location
OxPhos takes place at the inner mitochondrial membrane (IMM). This membrane is impermeable to protons, which is essential for maintaining the electrochemical gradient.
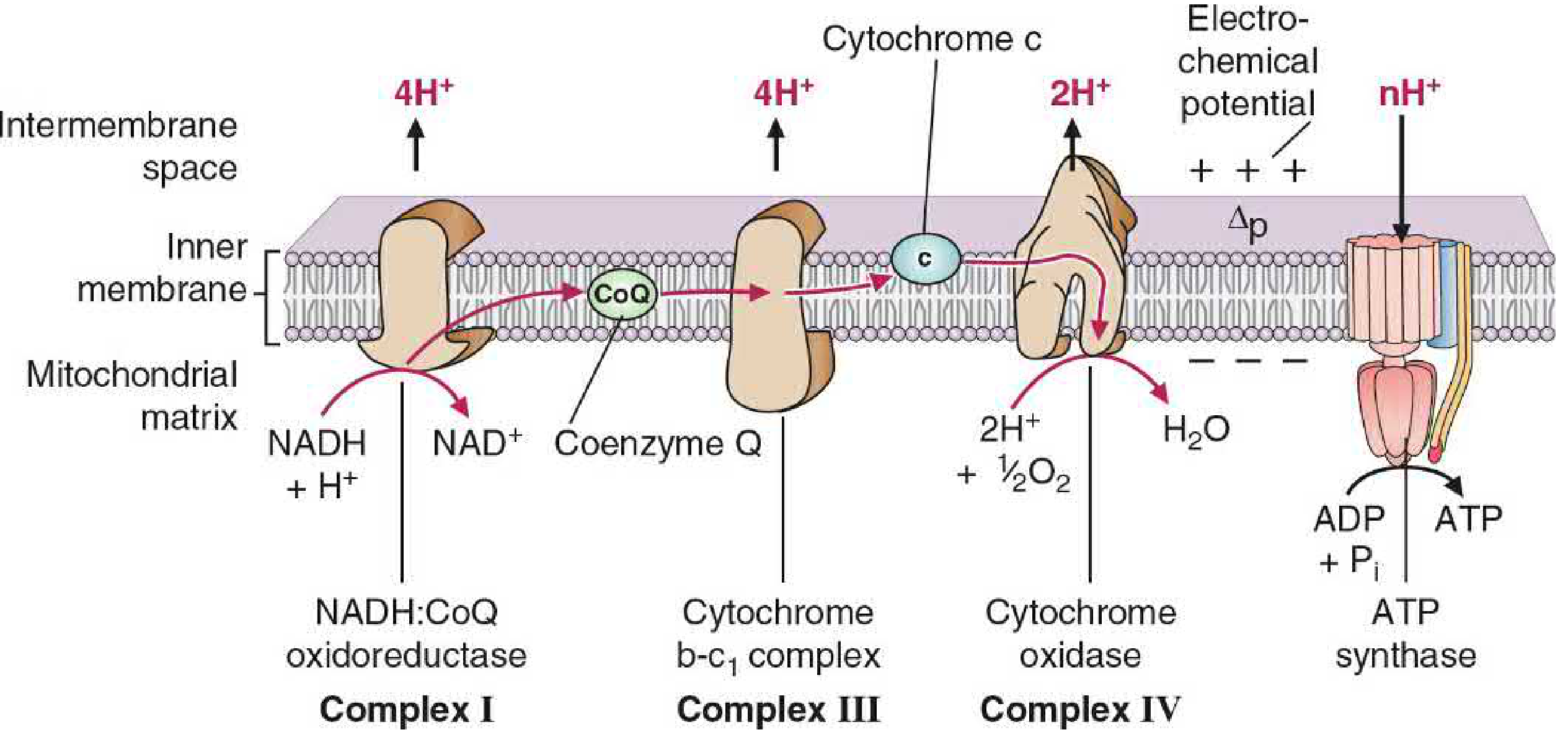
NADH donates electrons at Complex I; electrons travel via CoQ to Complex III, then cytochrome c to Complex IV. At each proton-pumping complex, H+ is moved from matrix to intermembrane space. The resulting gradient (Δp) drives ATP synthase.
The Electron Transport Chain (ETC)
Electrons flow through four multiprotein complexes embedded in the IMM:
| Complex | Name | Electron Carriers | Protons Pumped |
|---|---|---|---|
| I | NADH:CoQ oxidoreductase (NADH dehydrogenase) | FMN, Fe-S centers | 4 H+ |
| II | Succinate dehydrogenase | FAD, Fe-S centers | 0 H+ (no pumping) |
| III | Cytochrome b-c1 complex | Cytochrome b, Fe-S, cytochrome c1 | 4 H+ |
| IV | Cytochrome c oxidase | Cytochromes a, a3, Cu | 2 H+ |
Mobile carriers connecting the complexes:
- Coenzyme Q (CoQ / ubiquinone) - lipid-soluble, diffuses freely in the membrane; carries electrons from Complexes I and II to Complex III
- Cytochrome c - water-soluble protein in the intermembrane space; carries electrons from Complex III to Complex IV
At Complex IV, 4 electrons + 4H+ + O2 → 2 H2O. This is where cyanide and carbon monoxide inhibit (they bind Fe in cytochrome a3, blocking O2 reduction).
Chemiosmotic Model (Mitchell Hypothesis)
As electrons pass through Complexes I, III, and IV, protons are pumped from the matrix to the intermembrane space, generating:
- Membrane potential (ΔΨ) - electrical gradient (positive outside, negative inside)
- pH gradient (ΔpH) - chemical proton gradient
Together these form the proton-motive force (Δp):
Δp = ΔΨ + (−2.303 RT/F) × ΔpH
This electrochemical gradient is the immediate energy source for ATP synthesis.
ATP Synthase (Complex V / F0F1-ATPase)
ATP synthase has two domains:
- F0 (membrane domain): proton channel; contains the rotating c-ring subunits
- F1 (matrix headpiece): catalytic domain with three αβ subunit pairs
Binding change mechanism: As protons flow through F0, the asymmetric γ-shaft rotates, cycling each β subunit through three conformations - Open (binds ADP + Pi), Loose (ATP forms), and Tight (ATP releases). Three protons are translocated per ATP synthesized; one additional proton is consumed transporting phosphate into the matrix.
Regulation
OxPhos rate is tightly coupled to the rate of ATP utilization:
- When ATP is consumed → ADP rises → proton influx through ATP synthase accelerates → electron transport speeds up to restore Δp
- When ATP demand is low → Δp builds up (proton back-pressure) → electron transport slows
This is the respiratory control mechanism.
Inhibitors of OxPhos
| Agent | Site of Action | Mechanism |
|---|---|---|
| Rotenone, amobarbital | Complex I | Block NADH:CoQ oxidoreductase |
| Malonate | Complex II | Competitive inhibitor of succinate dehydrogenase |
| Antimycin A | Complex III | Blocks cytochrome b |
| Cyanide, CO, azide | Complex IV (cytochrome a3) | Bind Fe, prevent O2 reduction |
| Oligomycin | ATP synthase (F0) | Blocks proton channel |
| DNP (dinitrophenol) | IMM | Proton ionophore - uncoupler |
Uncoupling
Chemical uncouplers (proton ionophores) like DNP are lipid-soluble, carry protons across the IMM, and collapse the gradient. Electron transport continues (and even accelerates) but ATP synthesis stops; energy is dissipated as heat. DNP was once used as a weight-loss drug but caused fatal hyperthermia.
Physiological uncoupling occurs via:
- UCP1 (thermogenin) in brown adipose tissue - generates heat for thermogenesis in newborns and cold-adapted animals; activated by fatty acids, inhibited by purine nucleotides
- Other UCP isoforms (UCP2, UCP3) in other tissues with less well-defined roles
Energy Yield - Complete Glucose Oxidation
| Step | ATP Yield |
|---|---|
| Glycolysis (2 NADH via malate-aspartate shuttle) | 2 ATP + ~5 ATP |
| Pyruvate dehydrogenase (2 NADH) | ~5 ATP |
| TCA cycle (6 NADH + 2 FADH2 + 2 GTP) | ~20 ATP |
| Total per glucose | ~30-32 ATP |
To produce the same ATP per unit time using anaerobic glycolysis alone, glycolysis must run ~15 times faster.
NADH Shuttle Systems (Cytosolic NADH Entry)
The IMM is impermeable to NADH, so cytosolic NADH (from glycolysis) uses shuttles:
- Malate-aspartate shuttle - yields ~2.5 ATP per NADH; predominates in liver, heart, kidney
- Glycerol-3-phosphate shuttle - yields ~1.5 ATP per NADH (donates to FAD, not NAD); predominates in skeletal muscle and brain
Mitochondrial DNA and OXPHOS Diseases
- 13 of the ~90 ETC subunits are encoded by mtDNA (maternally inherited); the rest are nuclear-encoded
- Mutations in mtDNA or nuclear DNA cause OXPHOS diseases: lactic acidosis, myopathy, encephalopathy, retinal degeneration (tissues with high energy demand suffer most)
- Doxorubicin (anthracycline) cardiotoxicity involves binding cardiolipin, inhibiting succinate oxidation, inactivating cytochrome oxidase, and inhibiting ATP synthase
- AZT (zidovudine) can deplete mtDNA by inhibiting mtDNA polymerase γ, causing mitochondrial myopathy with ragged red fibers
Clinical Relevance
- Ischemia - O2 deprivation halts ETC, ATP falls, inner membrane permeability increases, mitochondria swell → irreversible cell injury and necrosis
- Cyanide poisoning - inhibits Complex IV; cells cannot use O2 despite normal hemoglobin function (histotoxic hypoxia)
- Thyroid hormone excess - increases UCPs and metabolic rate, increasing heat production
- Mitochondrial permeability transition pore (mPTP) - ANT + VDAC form a nonspecific channel under stress, triggering rapid necrotic cell death
Sources: Basic Medical Biochemistry - A Clinical Approach, 6e, pp. 862-884 | Biochemistry, 8th ed (Lippincott), Ch. 6
This is a shared conversation. Sign in to Orris to start your own chat.