MHC molecules and antigen presentation to T lymphocytes
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Reading File
Reading File
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Reading File
I now have all the content needed for a thorough answer. Let me compile the comprehensive response.
MHC Molecules and Antigen Presentation to T Lymphocytes
1. Discovery and Overview
The major histocompatibility complex (MHC) was originally discovered from studies of tissue graft rejection in mice. George Snell created congenic mouse strains that differed only at a single chromosomal locus - chromosome 17 in mice (called H-2). Grafts between strains that differed at this locus were rejected, giving the region its name: the major histocompatibility complex (histo = tissue). In humans, the equivalent proteins were identified as human leukocyte antigens (HLAs) - named because they were detected by antibodies in the serum of multiply transfused patients, multiparous women, and transplant recipients.
The fundamental immunological role of MHC molecules was clarified by Rolf Zinkernagel and Peter Doherty (Nobel Prize 1996), who demonstrated MHC restriction: virus-specific cytotoxic T lymphocytes (CTLs) only kill virus-infected cells if those cells share the same MHC molecules as the T cell's host. This showed that T cells recognize antigen and MHC simultaneously - a concept called dual recognition.
"The recognition of antigens by CD8+ CTLs is restricted by class I MHC molecules, and the responses of CD4+ helper T lymphocytes to antigens are restricted by class II MHC molecules." - Cellular and Molecular Immunology
2. MHC Genes
The MHC locus contains two main classes of polymorphic genes, plus nonpolymorphic genes involved in antigen processing:
| Gene Class | Location (human) | Product | Function |
|---|---|---|---|
| Class I | HLA-A, -B, -C | Transmembrane glycoprotein | Presents cytosolic peptides to CD8+ T cells |
| Class II | HLA-DR, -DP, -DQ | Heterodimer (α + β chains) | Presents endosomal peptides to CD4+ T cells |
| Class III | Between Class I & II | Complement proteins, cytokines (TNF, LT) | Innate/inflammatory functions |
Polymorphism is a hallmark of MHC genes - there are hundreds of alleles for each locus across the human population. A heterozygous individual expresses 6 different class I molecules (two each of HLA-A, -B, -C) and 8 or more class II molecules (multiple DR, DP, and DQ allotypes). This population-level diversity ensures that virtually any pathogen-derived peptide can be presented by some MHC allele in the species.
3. MHC Molecule Structure
Class I MHC
- Composed of a polymorphic α chain (3 extracellular domains: α1, α2, α3) non-covalently associated with the non-polymorphic β2-microglobulin (β2m)
- The peptide-binding groove is formed by the α1 and α2 domains (encoded by exons 2 and 3 of the class I gene)
- The groove is closed at both ends, restricting bound peptides to 8-11 amino acids
- Polymorphic residues cluster in and around the peptide-binding cleft, determining which peptides each allele can present
- Expressed on virtually all nucleated cells
Class II MHC
- A non-covalent heterodimer of α and β chains (both polymorphic in HLA-DR, -DP, -DQ)
- The peptide-binding groove is formed by the α1 domain (of α chain) + β1 domain (of β chain)
- The groove is open at both ends, allowing binding of longer peptides: 10-30+ residues (optimal 12-16)
- Expressed constitutively only on professional APCs: dendritic cells (DCs), B lymphocytes, macrophages, and thymic epithelium
- Upregulated on many cell types by IFN-γ
4. Antigen Processing Pathways
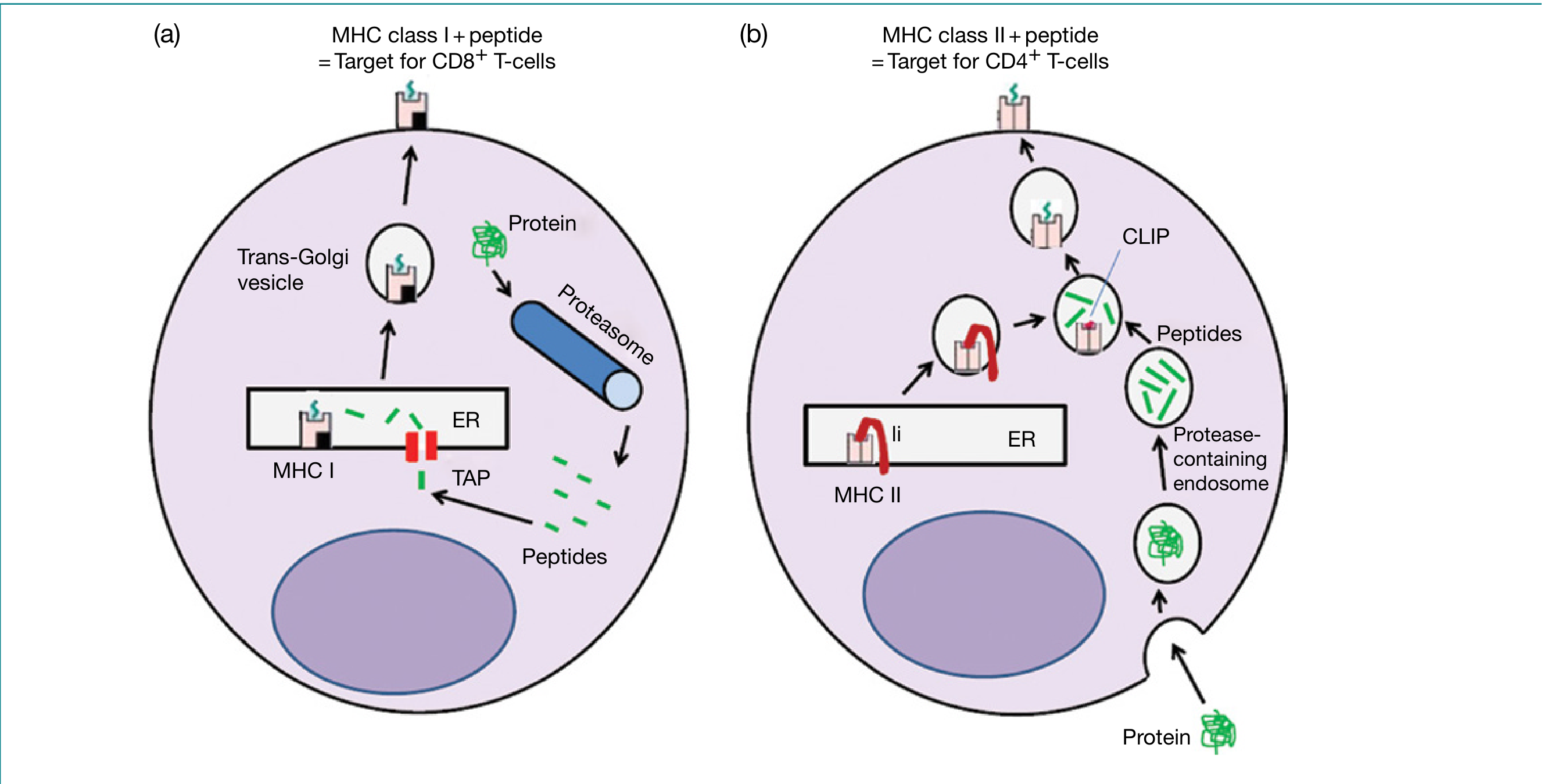
The two pathways are shown below (from Roitt's Essential Immunology):
And the detailed steps (from Cellular and Molecular Immunology):
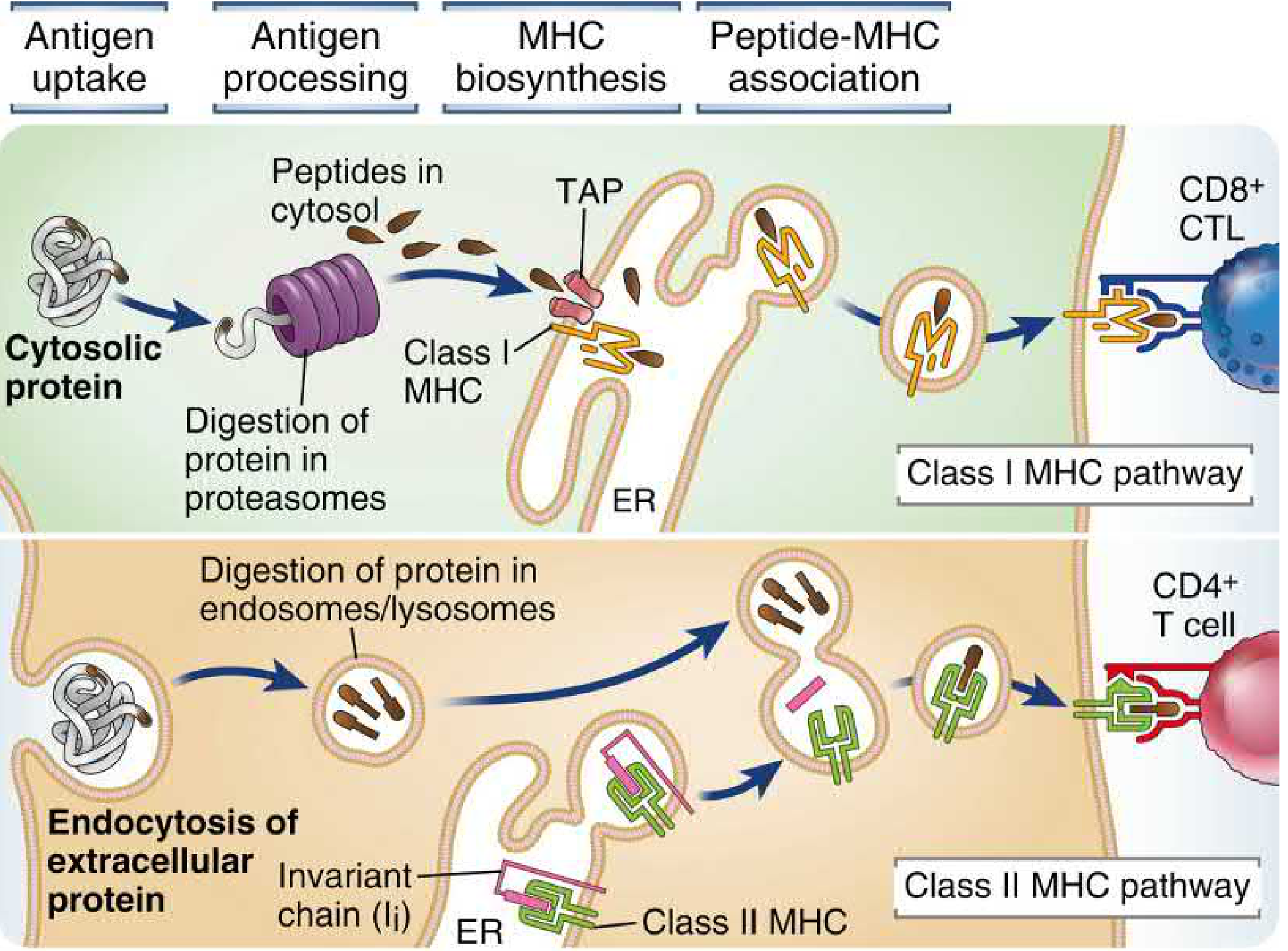
4a. Class I MHC Pathway (Endogenous / Cytosolic Antigens)
This pathway presents intracellular (cytosolic) proteins to CD8+ T cells.
Step 1 - Ubiquitination and proteasomal degradation
Cytosolic proteins targeted for degradation are tagged with polyubiquitin chains. The 26S proteasome (a barrel-shaped structure of 28 subunits in four stacked rings) degrades them into peptides. Viral proteins, tumor neoantigens, and misfolded ER proteins (via ER-associated degradation, ERAD) are all processed this way. IFN-γ replaces three standard proteasomal subunits (β1, β2, β5) with immunoproteasome subunits (LMP2, MECL-1, LMP7) that generate peptides with better C-terminal anchor residues for class I binding.
Step 2 - TAP transport into the ER
Peptides (typically 8-16 amino acids) are transported from the cytosol into the ER lumen by the TAP1/TAP2 heterodimer (Transporter Associated with antigen Processing) - ATP-dependent peptide pumps encoded within the MHC locus itself. Peptides that are too long can be trimmed by ER aminopeptidases (ERAP1, ERAP2 in humans; ERAAP in mice).
Step 3 - Peptide loading complex and MHC assembly
Inside the ER, nascent class I heavy chains assemble with β2m and are stabilized by the peptide loading complex (PLC), which includes: TAP1/2, tapasin (bridges TAP to class I), calreticulin (chaperone), and ERp57 (disulfide isomerase). An optimal peptide of 8-10 residues stabilizes the class I molecule and allows it to be released from the PLC.
Step 4 - Transport to cell surface
The peptide-MHC I complex travels through the Golgi to the plasma membrane, where it is displayed for inspection by CD8+ T cells.
4b. Class II MHC Pathway (Exogenous / Endosomal Antigens)
This pathway presents extracellular proteins internalized by endocytosis to CD4+ T cells.
Step 1 - Antigen uptake
Extracellular proteins are internalized by phagocytosis, macropinocytosis, or receptor-mediated endocytosis into early endosomes.
Step 2 - Endosomal processing
As early endosomes acidify and mature into late endosomes/lysosomes, proteases (cathepsins B, D, H, L, S, and asparagine endopeptidase) digest the internalized proteins into peptides 10-30 residues long. The enzyme GILT (IFN-γ-induced lysosomal thiol reductase) reduces disulfide bonds in engulfed proteins to facilitate unfolding and digestion.
Step 3 - Invariant chain and class II biosynthesis
In the ER, newly synthesized class II αβ heterodimers assemble with the invariant chain (Ii). Ii serves multiple roles:
- Acts as a dedicated chaperone ensuring correct folding
- Occupies the peptide-binding groove (via its CLIP segment - Class II-associated Invariant chain Peptide), blocking premature peptide loading in the ER
- Trimerizes to form a nonameric complex (3 αβ dimers + 3 Ii chains)
- Contains targeting motifs that direct the complex to the endocytic pathway
Step 4 - CLIP removal and HLA-DM editing
The class II-Ii complex is transported to the Golgi, then to MHC class II-enriched compartments (MIICs), where Ii is progressively degraded by cathepsins S/L and asparagine endopeptidase, leaving only the CLIP fragment in the groove. The non-classical MHC molecule HLA-DM (H-2M in mice) catalyzes the exchange of CLIP for high-affinity antigenic peptides. HLA-DO (H-2O) can inhibit HLA-DM activity, modulating the repertoire of presented peptides.
Step 5 - Surface display
Stable peptide-MHC II complexes travel to the plasma membrane. Surface class II levels are regulated by the ubiquitin E3 ligase MARCH-1, which targets class II for degradation; during infection, MARCH-1 is suppressed to increase surface antigen display.
5. Comparative Table: Class I vs. Class II Pathways
| Feature | Class I MHC | Class II MHC |
|---|---|---|
| T cell type activated | CD8+ CTLs | CD4+ helper T cells |
| Source of antigen | Cytosolic proteins (viral, tumor, ERAD) | Endosomal/lysosomal (extracellular antigens) |
| Site of degradation | Proteasome (cytosol) | Late endosomes/lysosomes |
| Proteolytic enzymes | Proteasomal subunits (β1, β2, β5; LMP2, LMP7, MECL-1) | Cathepsins B, D, H, L, S; AEP |
| Peptide transport | TAP1/TAP2 into ER | Ii chain guides class II to endosomes |
| Peptide size | 8-11 amino acids | 12-16 amino acids (up to 30+) |
| Chaperones / accessories | Tapasin, calreticulin, ERp57, calnexin | Invariant chain (Ii), HLA-DM/DO |
| APC expression | All nucleated cells | DCs, macrophages, B cells, thymic epithelium |
| Increased by | IFN-α, IFN-β, IFN-γ | IFN-γ (via CIITA) |
| Outcome | Killing of infected / tumor cells | Macrophage activation, B cell help, antibody production |
6. Cross-Presentation
A critical exception to the "two-pathway" rule is cross-presentation (cross-priming): specialized type 1 conventional dendritic cells (cDC1) can capture exogenous antigens (e.g., from virus-infected cells or tumor cells) and load them onto class I MHC molecules, thereby activating naive CD8+ T cells.
Mechanistically, antigens from phagocytosed cells or debris are taken into vesicles that fuse with ER membrane. By incompletely understood mechanisms, proteins translocate from vesicles into the cytosol, enter the standard proteasome-TAP-class I pathway, and are presented on class I. This allows the immune system to prime antitumor and antiviral CTL responses even when the antigen is not produced within a DC itself.
7. Expression and Regulation
- Class I expression: Constitutive on all nucleated cells; upregulated by IFN-α/β (during early innate antiviral response) and IFN-γ (during adaptive responses). This creates a positive feedback loop: viral infection triggers innate IFN production, which upregulates class I on all cells, making viral antigens more visible to CD8+ T cells.
- Class II expression: Constitutive on professional APCs only; transcriptionally activated by IFN-γ through the master regulator CIITA (class II transactivator). Many pathogens have evolved mechanisms to downregulate class II expression (e.g., herpes viruses blocking CIITA, Mycobacterium inhibiting phagolysosome maturation).
8. Physiological Significance
The segregation of antigen presentation into two pathways has profound functional logic:
- Cytosolic antigens (viruses, intracellular bacteria like Listeria, tumor neoantigens) are presented by class I MHC to CD8+ CTLs, which kill the infected/transformed cell - the only way to eliminate a pathogen producing protein from within a host cell.
- Extracellular antigens (bacteria, toxins, parasites, vaccine antigens) are presented by class II MHC to CD4+ T helper cells, which help macrophages kill phagocytosed microbes and help B cells make antibodies - the appropriate defenses against extracellular threats.
This division ensures that the antigen receptor of a T cell (which cannot by itself distinguish where an antigen comes from) is linked to the correct effector function by the MHC molecule type that co-presents the antigen.
MHC molecules also determine immunodominance: of the many peptides generated by proteolysis of a complex antigen, only those that bind most avidly to the individual's MHC alleles will be efficiently presented. These become the immunodominant epitopes, which has direct applications in vaccine design - synthetic peptides containing such epitopes can serve as candidate vaccines.
9. Non-Classical MHC and CD1
Beyond classical class I and II, there are:
- Non-classical class I molecules (HLA-E, HLA-F, HLA-G): limited polymorphism, specialized roles (e.g., HLA-E presents signal peptides to NK receptors; HLA-G mediates maternal-fetal tolerance)
- CD1 molecules: MHC-like but present lipid antigens (e.g., mycobacterial lipoarabinomannan) to NKT cells and certain T cells
- MR1: presents vitamin B metabolites (riboflavin precursors) to mucosal-associated invariant T (MAIT) cells
Sources:
- Cellular and Molecular Immunology (Abbas, Lichtman, Pillai), Chapter 6
- Roitt's Essential Immunology, Chapter 5
- Janeway's Immunobiology 10e, Chapter 6
This is a shared conversation. Sign in to Orris to start your own chat.