Name of ketone bodies Describe formation and utilisation of ketone bodies
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
I now have everything needed for a thorough, well-illustrated answer. Let me compose it.
Ketone Bodies
Names of the Three Ketone Bodies
| Ketone Body | Also Known As | Notes |
|---|---|---|
| Acetoacetate | Acetoacetic acid | Primary ketone body; the hub of ketone metabolism |
| Beta-hydroxybutyrate (β-hydroxybutyrate) | D-3-hydroxybutyrate | Quantitatively the most abundant in blood/urine during ketosis; strictly not a "ketone" chemically (it has a hydroxyl group, not a keto group), but classified as a ketone body |
| Acetone | Dimethyl ketone | Formed by spontaneous, non-enzymatic decarboxylation of acetoacetate; volatile, excreted via lungs |
The interrelationship of these three is shown below:
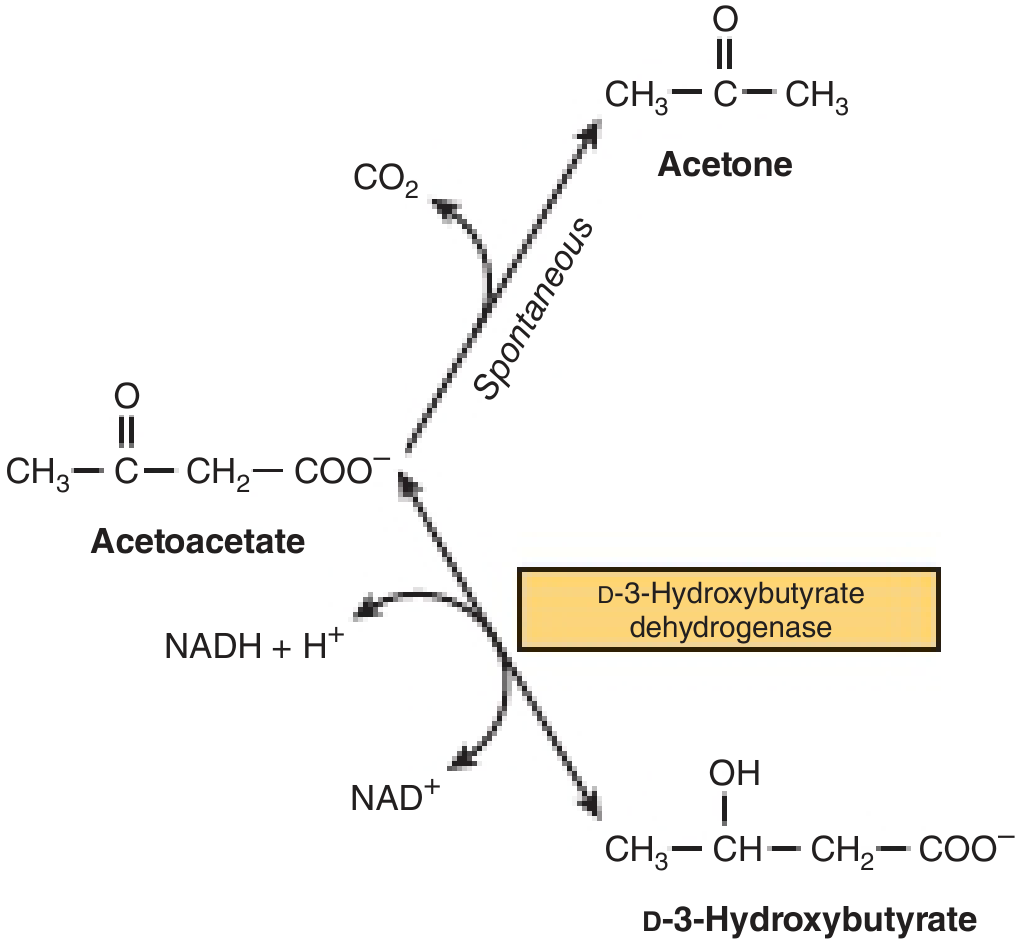
Formation of Ketone Bodies (Ketogenesis)
Site: Exclusively in the hepatic mitochondrial matrix (and rumen epithelium in ruminants). Extrahepatic tissues lack the necessary enzymes (particularly mitochondrial HMG-CoA synthase) to produce ketone bodies.
Conditions that trigger ketogenesis:
- Prolonged fasting / starvation
- Uncontrolled diabetes mellitus (low insulin : glucagon ratio)
- High-fat, low-carbohydrate diet
- Prolonged heavy exercise
Step-by-Step Pathway
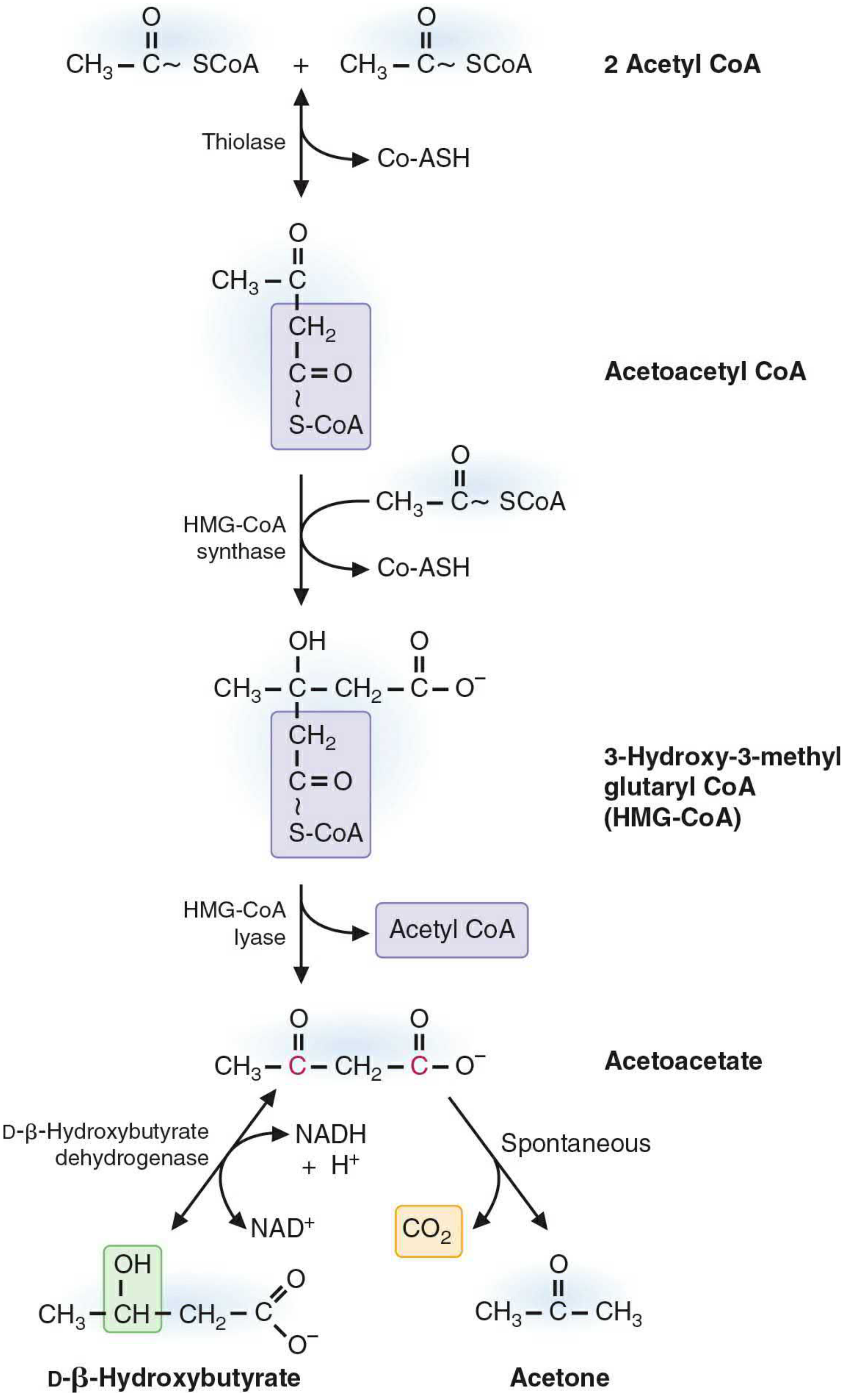
Step 1 - Acetoacetyl-CoA formation
Two molecules of acetyl-CoA (generated from beta-oxidation of fatty acids) condense via thiolase (reversal of the beta-oxidation thiolase reaction) to form acetoacetyl-CoA, releasing one CoA-SH.
Step 2 - HMG-CoA formation
Acetoacetyl-CoA condenses with another acetyl-CoA via HMG-CoA synthase (the rate-limiting, key regulatory enzyme of ketogenesis) to form 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA), releasing CoA-SH.
Step 3 - Acetoacetate release
HMG-CoA lyase cleaves HMG-CoA, releasing acetyl-CoA and free acetoacetate. Both HMG-CoA synthase and HMG-CoA lyase must be present in the mitochondria for ketogenesis to occur.
Step 4 - From acetoacetate, two fates:
- Reduction to beta-hydroxybutyrate: acetoacetate is reduced by mitochondrial D-3-hydroxybutyrate dehydrogenase using NADH + H⁺ → NAD⁺. The equilibrium between the two is controlled by the NAD⁺/NADH redox state of the mitochondria. During heavy beta-oxidation, NADH is abundant, so the ratio of beta-hydroxybutyrate : acetoacetate shifts toward ~3:1.
- Spontaneous decarboxylation to acetone: non-enzymatic, losing CO₂. Acetone is excreted via the lungs.
Regulation of Ketogenesis
- Increased free fatty acid supply from adipose tissue (lipolysis activated by glucagon/low insulin)
- Decreased insulin inactivates acetyl-CoA carboxylase → decreased malonyl-CoA → releases inhibition of CPT-1 (carnitine palmitoyltransferase I) → fatty acyl-CoA enters mitochondria for beta-oxidation
- High NADH/NAD⁺ ratio inhibits the TCA cycle (oxaloacetate is diverted to malate/gluconeogenesis), so acetyl-CoA cannot enter the TCA cycle and is redirected to ketogenesis
- Increased transcription of mitochondrial HMG-CoA synthase gene during prolonged fasting
Utilisation of Ketone Bodies
Site: All extrahepatic tissues (brain, heart, skeletal muscle, renal cortex, intestinal mucosa, etc.) except liver and red blood cells.
The liver produces ketone bodies but cannot utilize them - it lacks succinyl-CoA transferase (CoA transferase / thiophorase), the key enzyme for ketone reactivation. This ensures a net outflow of ketone bodies from liver to peripheral tissues.
The overall flow is illustrated below:
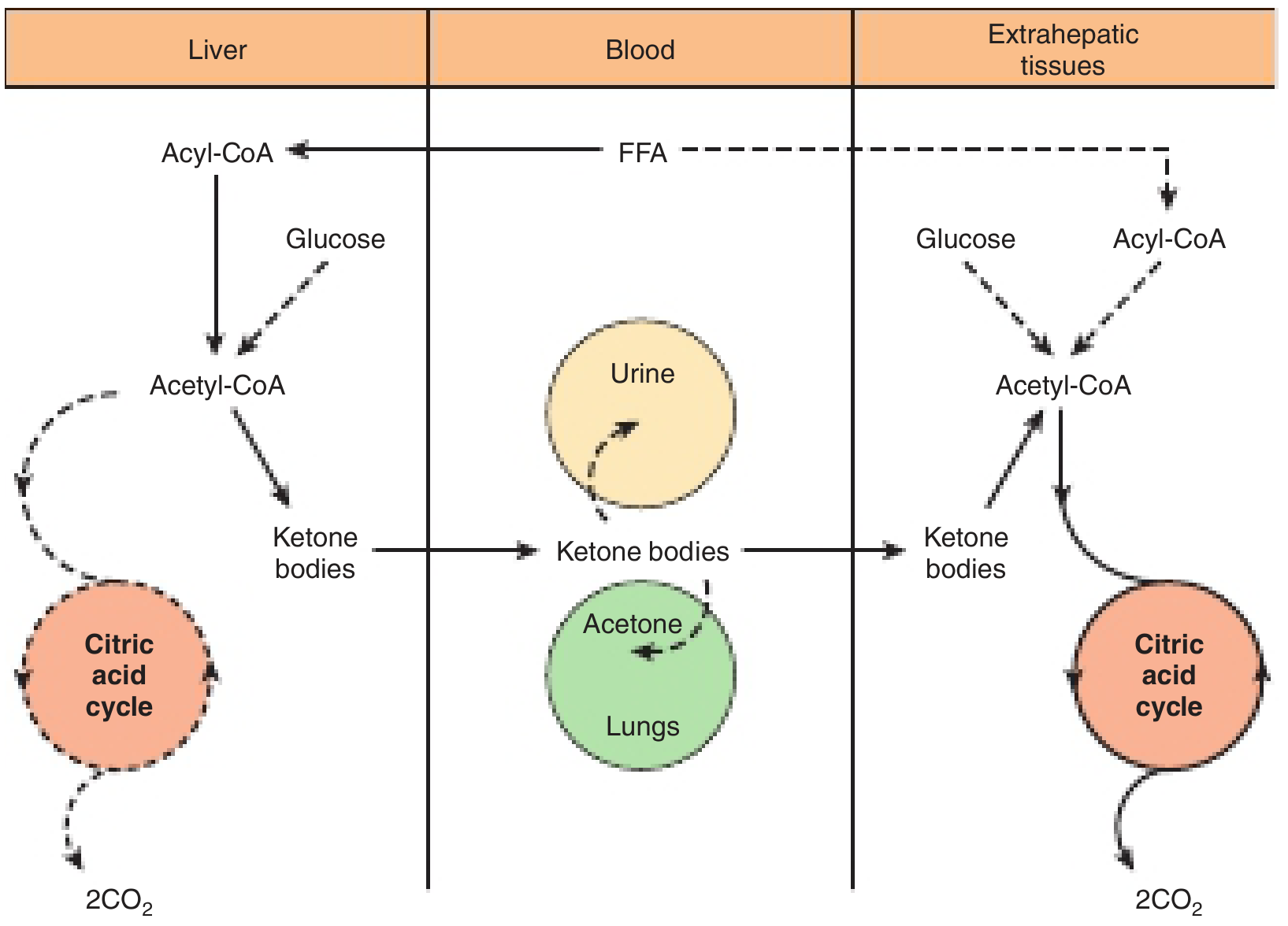
Step-by-Step Utilization Pathway
Step 1 - Beta-hydroxybutyrate → Acetoacetate
In extrahepatic tissue mitochondria, beta-hydroxybutyrate is oxidized back to acetoacetate by D-3-hydroxybutyrate dehydrogenase, producing NADH.
Step 2 - Reactivation of acetoacetate (key step)
Acetoacetate receives CoA from succinyl-CoA via succinyl-CoA-acetoacetate-CoA transferase (thiophorase). This yields acetoacetyl-CoA and succinate. This reaction is absent in the liver (ensuring net ketone body export).
Step 3 - Thiolase cleavage
Acetoacetyl-CoA is cleaved by thiolase (3-ketothiolase) to yield 2 molecules of acetyl-CoA.
Step 4 - TCA cycle oxidation
The two acetyl-CoA molecules enter the citric acid cycle and are oxidized to CO₂ and H₂O, generating ATP.
ATP yield:
- 1 mol acetoacetate → ~19 mol ATP
- 1 mol beta-hydroxybutyrate → ~21.5 mol ATP (extra NADH from the dehydrogenase step)
Tissues and Context
| Tissue | Ketone Body Use |
|---|---|
| Brain | Uses extensively during prolonged starvation (replaces ~2/3 of glucose needs) |
| Skeletal muscle | Active user during fasting and exercise |
| Heart | Preferential fuel; uses ketones over glucose when available |
| Renal cortex | Major ketone consumer |
| Intestinal mucosa | Uses ketones + amino acids (not fatty acids) during starvation |
| Fetus | Ketone bodies cross the placenta and are used as fuel |
| Adipocytes | Can use ketones but not fatty acids during fasting |
| Liver | Produces but cannot utilize ketone bodies |
| RBCs | Cannot use (lack mitochondria) |
Normal Blood Levels and Clinical Significance
| State | Total ketone body concentration |
|---|---|
| Well-fed | < 0.2 mmol/L |
| Prolonged fasting | 4-5 mmol/L |
| Ketoacidosis threshold | > 7 mmol/L (exceeds bicarbonate buffer, causes acidosis and Kussmaul respiration) |
| Oxidative machinery saturation | ~12 mmol/L |
During uncontrolled type 1 diabetes, the relative excess of glucagon (a potent stimulator of ketogenesis) combined with increased fatty acid supply markedly elevates ketone body formation. Utilization fails to keep pace, leading to diabetic ketoacidosis (DKA).
Sources: Harper's Illustrated Biochemistry, 32nd Ed, pp. 232-233; Basic Medical Biochemistry - A Clinical Approach, 6th Ed, pp. 1097-1103
This is a shared conversation. Sign in to Orris to start your own chat.