Classify phospholipids with examples. Mention their specific role in maintaining the fluidity of plasma membrane
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
I now have comprehensive information from multiple authoritative textbooks. Let me compose the full answer.
Classification of Phospholipids
Phospholipids are the predominant lipids of cell membranes. They are amphipathic molecules - each has a hydrophilic (polar) head group and hydrophobic (nonpolar) fatty acid tails. Based on their backbone structure, they are divided into two major classes:
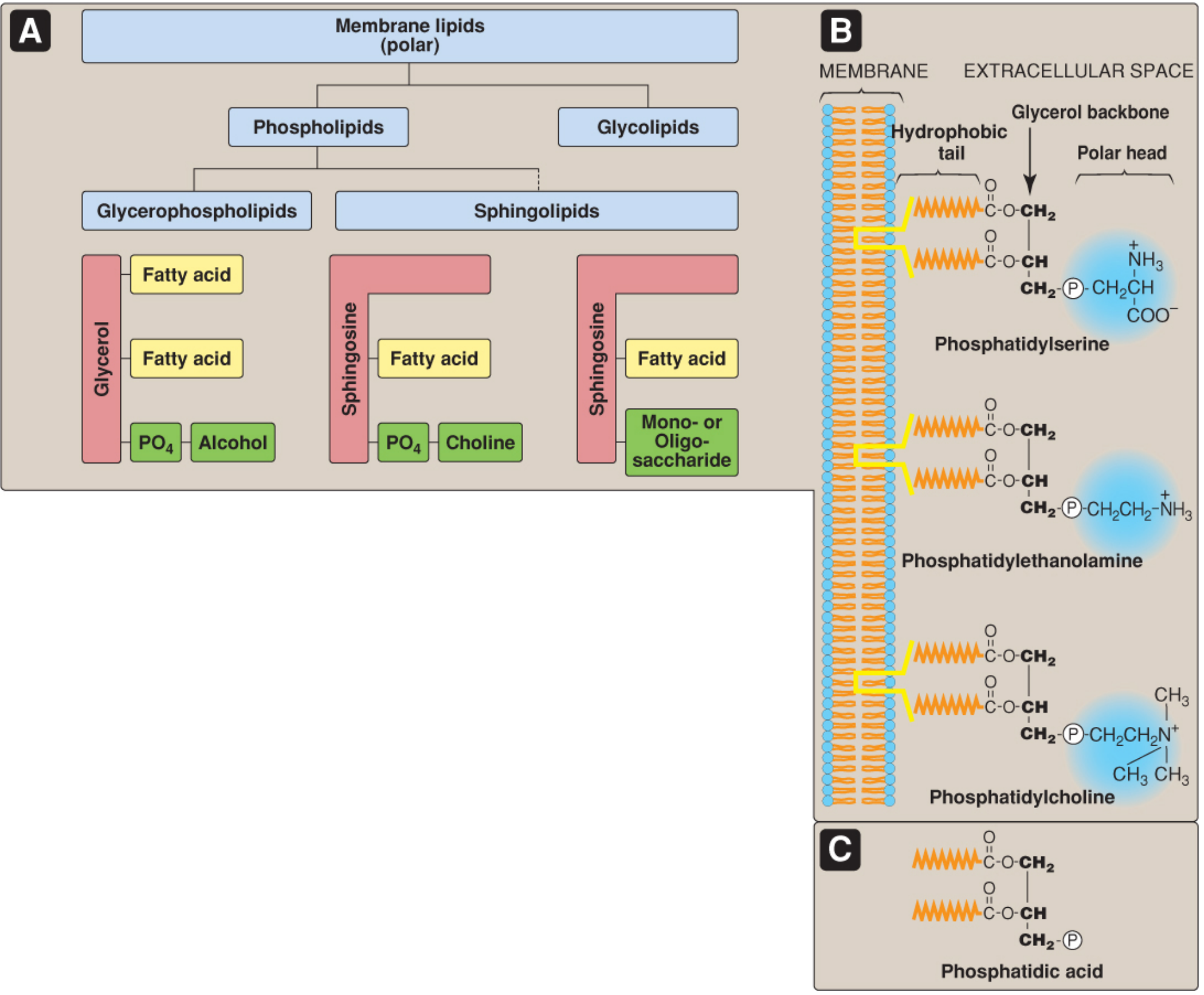
Class 1: Glycerophospholipids (Glycerophosphoglycerides)
These have a glycerol backbone and constitute the major class of membrane phospholipids. All are derived from (or are derivatives of) phosphatidic acid (PA) - diacylglycerol (DAG) with a phosphate group at carbon 3.
The phosphate on PA is esterified to an alcohol head group. This determines the specific glycerophospholipid formed:
| Alcohol head group | Glycerophospholipid formed | Notable role |
|---|---|---|
| Serine | Phosphatidylserine (PS) | Found on inner leaflet; exposed during apoptosis (signals phagocytosis) |
| Ethanolamine | Phosphatidylethanolamine (PE) | Abundant in nerve tissue; inner leaflet |
| Choline | Phosphatidylcholine (PC) / Lecithin | Most abundant phospholipid in most membranes; outer leaflet; component of lung surfactant and bile |
| Inositol | Phosphatidylinositol (PI) | Minor in quantity but major in signaling - PI(4,5)P2 is cleaved by phospholipase C to generate IP3 and DAG (second messengers) |
| Glycerol | Phosphatidylglycerol (PG) | Precursor to cardiolipin; found in lung surfactant |
Special Glycerophospholipids
-
Cardiolipin (diphosphatidylglycerol): Two PA molecules esterified to a central glycerol. Found almost exclusively in the inner mitochondrial membrane, where it stabilizes respiratory chain complexes. It is also clinically relevant - antibodies against cardiolipin form in syphilis (basis of the Wassermann test).
-
Plasmalogens (Ether phospholipids): The fatty acid at C-1 is replaced by an unsaturated alcohol attached via an ether bond (rather than an ester bond). Examples include plasmenylcholine (abundant in heart muscle) and plasmenylethanolamine (abundant in nerve tissue). Plasmalogens constitute ~18% of phospholipids in humans and are especially prevalent in brain and heart.
-
Platelet-activating factor (PAF): An ether lipid with an acetyl group at C-2 instead of a long fatty acid chain. Potent mediator of inflammation and platelet aggregation.
Class 2: Sphingophospholipids
These have a sphingosine backbone (derived from serine and palmitoyl-CoA) instead of glycerol. The only major sphingophospholipid is:
- Sphingomyelin: Ceramide (sphingosine + fatty acid) with phosphocholine as the head group. It is the only phospholipid not derived from glycerol. Sphingomyelin is abundant in the myelin sheath of neurons and the outer leaflet of plasma membranes. It is degraded by sphingomyelinase; deficiency causes Niemann-Pick disease.
Summary Classification Tree
Phospholipids
├── Glycerophospholipids (glycerol backbone)
│ ├── Phosphatidylcholine (PC) — "Lecithin"
│ ├── Phosphatidylethanolamine (PE) — "Cephalin"
│ ├── Phosphatidylserine (PS)
│ ├── Phosphatidylinositol (PI) and its phosphorylated forms
│ ├── Phosphatidylglycerol (PG)
│ ├── Cardiolipin (diphosphatidylglycerol)
│ └── Plasmalogens / Ether lipids (PAF, plasmenylcholine)
└── Sphingophospholipids (sphingosine backbone)
└── Sphingomyelin
Role of Phospholipids in Maintaining Plasma Membrane Fluidity
Membrane fluidity refers to the viscosity of the lipid bilayer - specifically, how readily phospholipid and protein molecules can move laterally within the plane of the membrane. Phospholipids determine fluidity through several mechanisms:
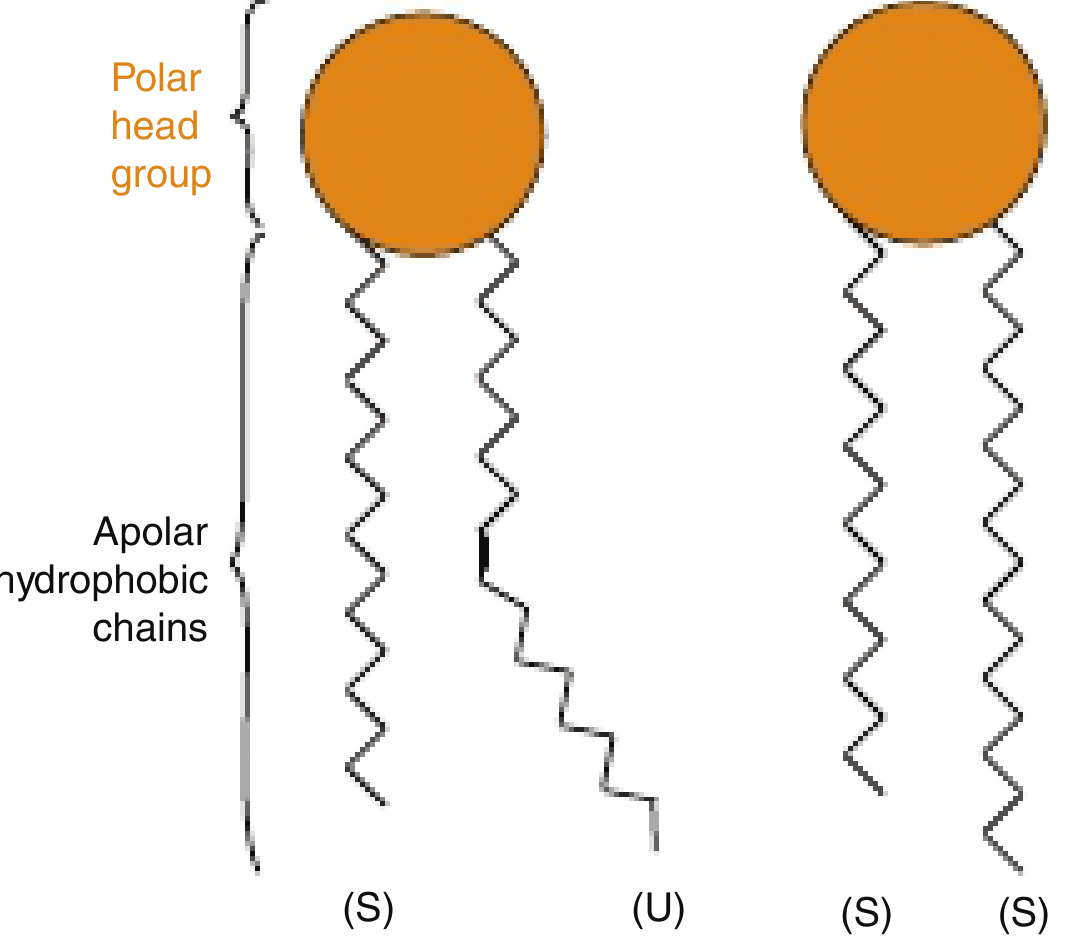
1. Fatty Acid Chain Length
Longer fatty acid chains have more surface area for van der Waals interactions with neighboring chains, causing them to pack more tightly together. This raises the transition temperature (Tm) - the temperature at which the membrane shifts from an ordered (gel-like/crystalline) phase to a disordered (liquid-like/fluid) phase. Shorter chains reduce these interactions and lower Tm, making the membrane more fluid at a given temperature.
2. Degree of Unsaturation (Most Important Factor)
- Saturated fatty acids form straight, extended chains that pack tightly and in parallel, creating a relatively stiff, ordered bilayer.
- Unsaturated fatty acids (in the cis configuration, as they are in biological membranes) introduce a kink in the hydrocarbon chain at each double bond. These kinks prevent tight packing of adjacent chains and increase the spacing between molecules.
- The result: more double bonds = more kinks = less packing = greater fluidity and lower Tm.
- Phospholipids of cellular membranes generally contain at least one unsaturated fatty acid with at least one cis double bond, which keeps the membrane in a fluid state at physiological temperature (37°C).
- Trans fatty acids, by contrast, do not create kinks and behave more like saturated chains - their incorporation into membranes reduces fluidity, which is one reason dietary trans fats are harmful.
3. Phospholipid Head Group Composition
The polar head group also influences packing. Smaller head groups (like PE) allow closer packing of the hydrophobic tails. Larger or more charged head groups (like PC with its bulky choline) create steric repulsion that loosens packing and increases fluidity.
4. Asymmetric Distribution Across Leaflets
Phospholipids are asymmetrically distributed between the two leaflets of the plasma membrane:
- Outer leaflet: PC and sphingomyelin (more saturated, tightly packed, less fluid)
- Inner (cytoplasmic) leaflet: PE and PS (more unsaturated, more fluid)
This asymmetry is actively maintained by flippases (ATP-dependent, move PS and PE from outer to inner leaflet) and floppases (move PC from inner to outer leaflet), and scramblases (bidirectional, calcium-activated; used in apoptosis signaling).
5. Role of Cholesterol as a Fluidity Buffer
While not a phospholipid, cholesterol works in concert with phospholipids. It inserts between phospholipid tails with its rigid steroid ring:
- Below Tm: Disrupts tight packing of saturated tails → increases fluidity
- Above Tm: Restricts the movement of unsaturated tails → decreases fluidity
This "buffering" effect keeps membrane fluidity within a narrow, physiologically optimal range regardless of temperature.
6. Biological Significance of Membrane Fluidity
As noted in Harper's Biochemistry: "As membrane fluidity increases, so does its permeability to water and other small hydrophilic molecules. The lateral mobility of integral proteins increases as the fluidity of the membrane increases." Insulin receptor binding efficiency, for instance, improves as unsaturated fatty acid content and fluidity increase. Bacteria dynamically remodel their phospholipid composition in response to temperature changes to maintain optimal membrane fluidity - a phenomenon called homeoviscous adaptation.
Sources:
- Lippincott's Illustrated Reviews: Biochemistry, 8th Ed, pp. 566-570 (Chapter 17: Phospholipid, Glycosphingolipid, and Eicosanoid Metabolism)
- Harper's Illustrated Biochemistry, 32nd Ed, pp. 481-485 (Chapter 40: The Fluid Mosaic Model)
- Junqueira's Basic Histology, 17th Ed, pp. 72-74 (Chapter 2: The Plasma Membrane)
This is a shared conversation. Sign in to Orris to start your own chat.