neuro muscular junction from human physiology sembulingam
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Reading File
Loading Image
Reading File
Loading Image
Reading File
Loading Image
I now have all the material needed for a comprehensive, well-illustrated answer on the neuromuscular junction. Let me compile it.
Neuromuscular Junction (NMJ)
Content synthesized from Ganong's Review of Medical Physiology (26th ed.) and Guyton & Hall Textbook of Medical Physiology - the standard reference physiology texts covering the same material as Sembulingam's Human Physiology.
Definition
The neuromuscular junction (NMJ) - also called the myoneural junction - is the specialized synapse where a motor nerve fiber (alpha motor neuron) terminates on a skeletal muscle fiber. It is the site of neuromuscular transmission, converting an electrical nerve impulse into a chemical signal that triggers muscle contraction.
Structure of the NMJ
The NMJ has three parts: the presynaptic terminal, the synaptic cleft, and the postsynaptic (motor end plate) membrane.
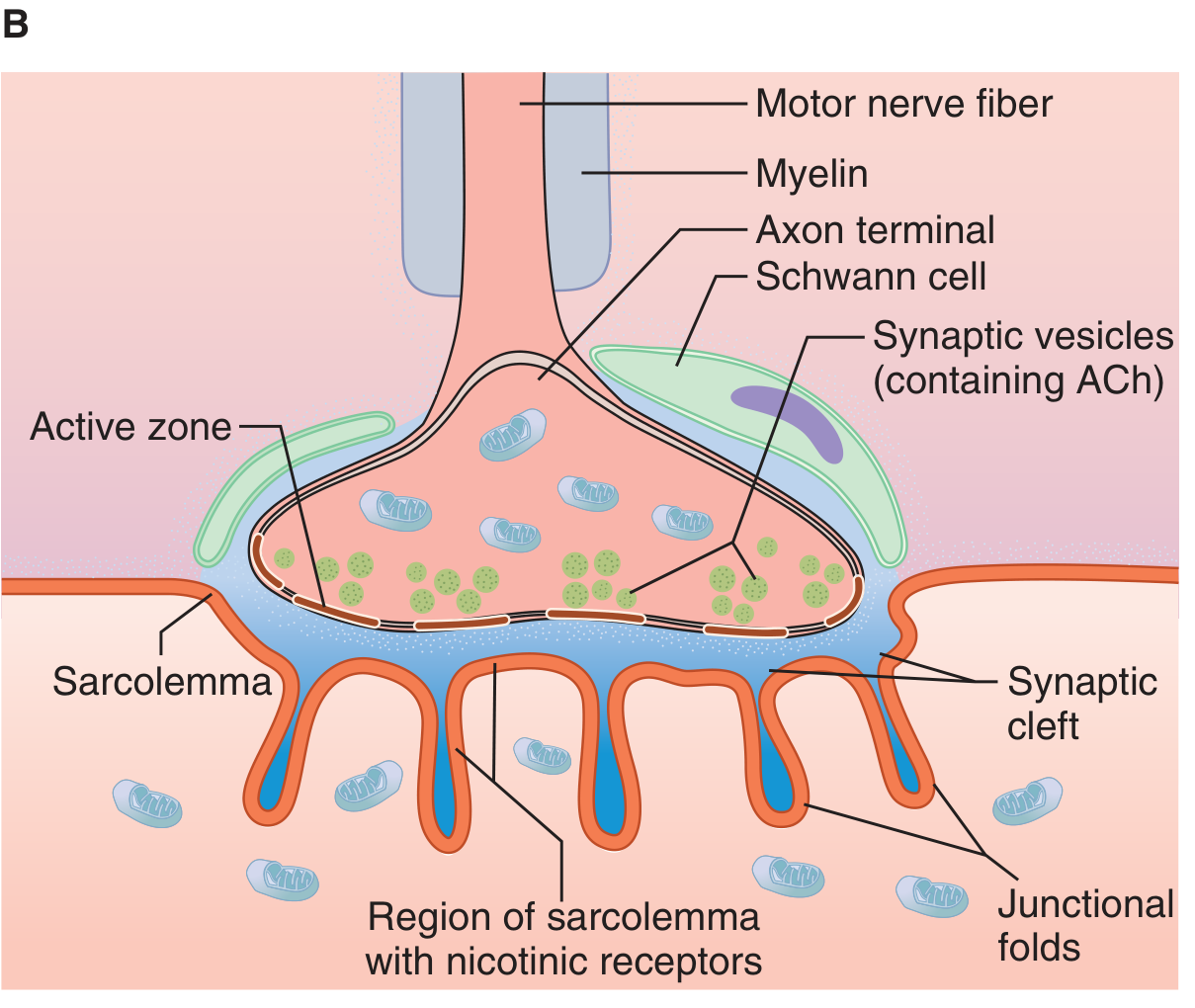
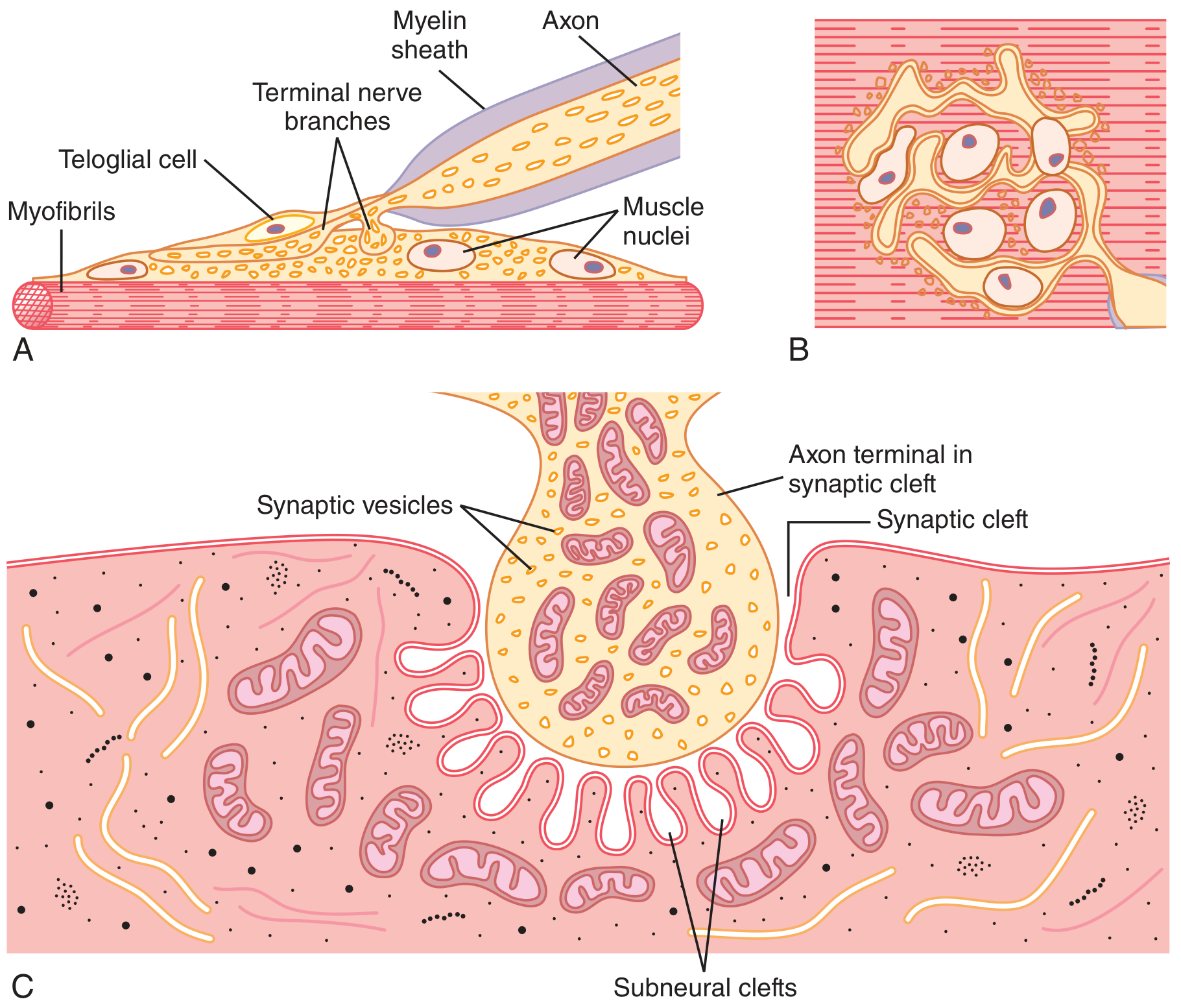
1. Presynaptic Terminal (Axon Terminal / Terminal Bouton)
- The motor nerve fiber loses its myelin sheath as it approaches the muscle fiber and divides into multiple terminal boutons
- Each terminal bouton contains:
- Numerous mitochondria (supply ATP for ACh synthesis)
- ~300,000 synaptic vesicles, each containing ~10,000 molecules of acetylcholine (ACh)
- Voltage-gated Ca²+ channels flanking dense bars on the inner membrane surface (active zones)
- The terminal is covered by Schwann cells (teloglial cells) that insulate it from surrounding fluids
2. Synaptic Cleft
- A gap of 20-30 nanometers between the nerve terminal and the muscle membrane
- Contains acetylcholinesterase (AChE) embedded in a fine connective tissue sponge - this enzyme rapidly destroys ACh after it acts
3. Postsynaptic Membrane (Motor End Plate)
- The thickened, specialized portion of the muscle (sarcolemma) beneath the terminal
- Forms deep junctional folds (subneural clefts) that greatly increase surface area
- The mouths of the junctional folds are densely packed with nicotinic cholinergic (N-M) receptors
- Voltage-gated Na+ channels line the depths of the subneural clefts
- Each endplate receives input from only one nerve fiber
ACh Synthesis and Storage
- Vesicles (~40 nm) are formed in the Golgi apparatus of the motor neuron cell body in the spinal cord
- They are transported via axoplasmic streaming down the axon to the terminal
- ACh is synthesized in the cytosol of the terminal:
- Choline + Acetyl-CoA → ACh (catalyzed by choline acetyltransferase)
- ACh is actively transported into vesicles and stored (~10,000 molecules/vesicle)
- ~300,000 vesicles are available in a single end plate
Sequence of Events in Neuromuscular Transmission
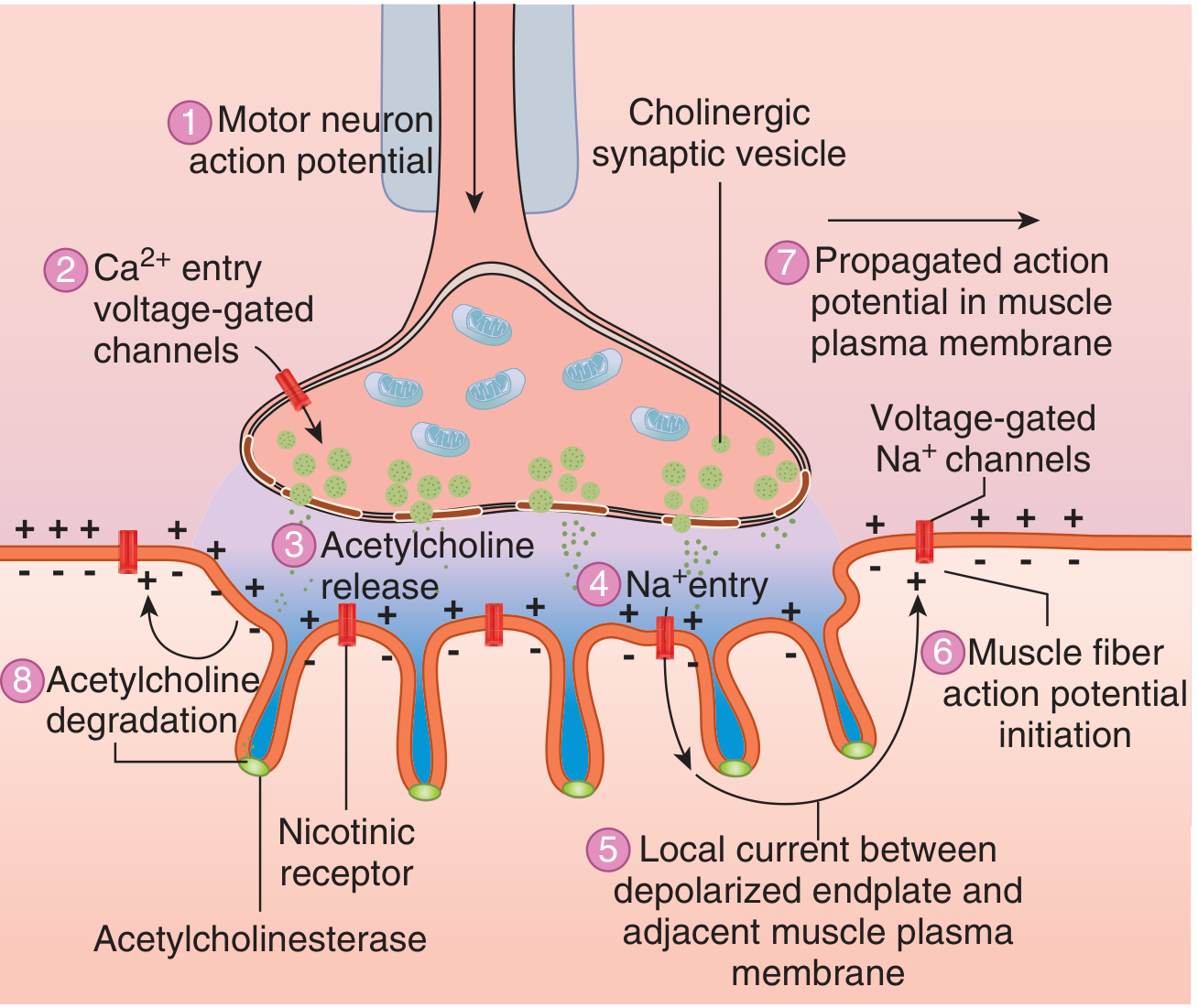
| Step | Event |
|---|---|
| 1 | Action potential (AP) arrives at axon terminal |
| 2 | Voltage-gated Ca²+ channels open; Ca²+ enters terminal (~100-fold increase in [Ca²+]) |
| 3 | Ca²+ activates calmodulin-dependent kinase → phosphorylates synapsin proteins → vesicles detach from cytoskeleton, migrate to active zone, and undergo exocytosis (~125 vesicles released per AP) |
| 4 | ACh diffuses across the synaptic cleft (20-30 nm) and binds to nicotinic N-M receptors on the motor end plate |
| 5 | Each receptor requires 2 ACh molecules to bind (one per α-subunit) → conformational change opens the ion channel |
| 6 | Na+ floods in; K+ moves out (Na+ influx >> K+ efflux) → end plate potential (EPP) of 50-75 mV is generated |
| 7 | EPP depolarizes adjacent sarcolemma beyond threshold → opens voltage-gated Na+ channels → action potential generated on both sides of the endplate |
| 8 | AP propagates in both directions along the muscle fiber → muscle contracts |
| 9 | ACh is destroyed by acetylcholinesterase in the synaptic cleft within a few milliseconds; choline is reabsorbed into terminal for re-synthesis |
The Nicotinic ACh Receptor
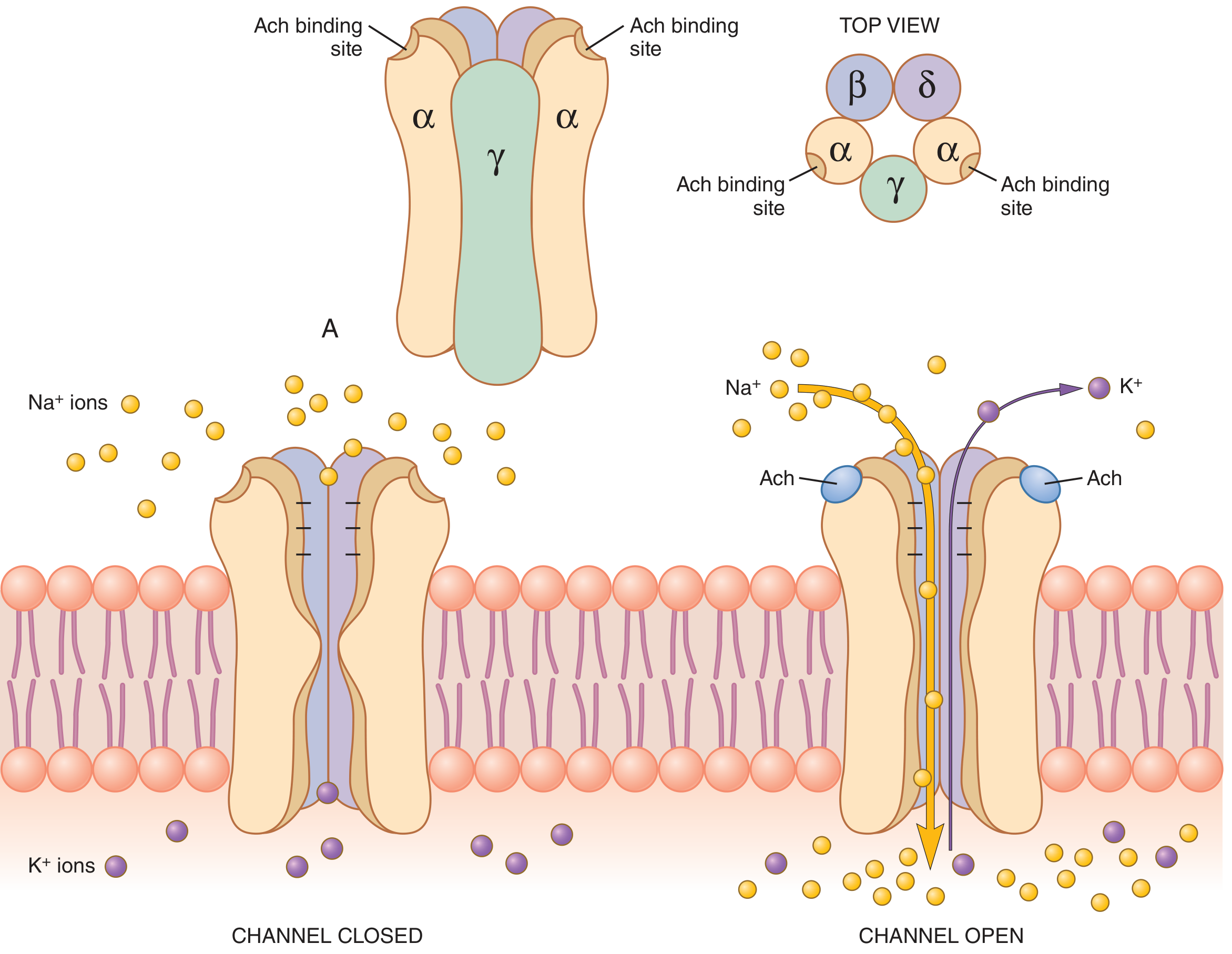
- Pentameric protein complex (MW ~275,000)
- Subunits: 2α + β + δ + γ (fetal) or 2α + β + δ + ε (adult)
- Five subunits surround a central ligand-gated ion channel
- ACh binds to the two α-subunits; both must bind for channel to open
- Channel diameter: ~0.65 nm - allows Na+, K+, Ca²+ to pass but not Cl- (negative charges at channel mouth repel anions)
- Each open channel can transmit 15,000-30,000 Na+ ions per millisecond
End Plate Potential (EPP)
- Na+ entry creates a local depolarization: the end plate potential
- EPP is normally ~50-75 mV (threshold for AP is only ~20-30 mV above resting)
- This gives a safety factor of ~3x - the EPP is normally 3 times larger than required to trigger an AP
- Weakness of EPP occurs with: curare (blocks ACh receptor) or botulinum toxin (blocks ACh release)
Quantal Release of ACh
- ACh is released in discrete packages called quanta (one quantum = contents of one vesicle)
- At rest: spontaneous release of single quanta produces tiny depolarizations called miniature end plate potentials (MEPPs) - ~0.5 mV each
- With AP: ~60-125 quanta released simultaneously → large EPP exceeding threshold
- Quantal size is proportional to [Ca²+] and inversely proportional to [Mg²+]
Recycling of Vesicle Membrane
- After exocytosis, clathrin-coated pits form in the terminal membrane within seconds
- These pinch off to form new vesicles within ~20 seconds
- New ACh is transported into the recycled vesicles - ready for the next impulse
Drugs Acting at the NMJ
| Drug / Agent | Mechanism | Effect |
|---|---|---|
| Curare (d-tubocurarine) | Competitive block of nicotinic receptors | Muscle paralysis (non-depolarizing) |
| Succinylcholine | Persistent depolarization (acts like ACh but not hydrolyzed quickly) | Depolarizing block |
| Neostigmine, Physostigmine | Inhibit acetylcholinesterase | Increased ACh, prolonged EPP (used in myasthenia gravis) |
| Methacholine, Carbachol | ACh-like agonists, resistant to AChE | Prolonged depolarization, muscle spasm |
| Botulinum toxin | Blocks ACh release (cleaves SNARE proteins) | Flaccid paralysis |
| Organophosphates (DFP) | Irreversibly inhibit AChE | Excess ACh accumulation |
Clinical Disorders of the NMJ
| Condition | Mechanism |
|---|---|
| Myasthenia Gravis | Autoimmune destruction/blockade of nicotinic ACh receptors → reduced EPP → fatigable muscle weakness |
| Lambert-Eaton Myasthenic Syndrome | Autoantibodies against voltage-gated Ca²+ channels in presynaptic terminal → reduced ACh release |
| Botulism | Clostridium botulinum toxin cleaves SNARE proteins → prevents vesicle docking/fusion → flaccid paralysis |
Safety Factor
Under normal conditions, each nerve impulse generates an EPP approximately 3 times larger than needed to trigger a muscle AP. This large safety margin ensures reliable neuromuscular transmission. Fatigue occurs only during extremely high-frequency stimulation (>100 Hz for several minutes), when ACh vesicle stores become temporarily depleted.
Sources: Ganong's Review of Medical Physiology 26th ed., p. 141 | Guyton & Hall Textbook of Medical Physiology, pp. 104-108
This is a shared conversation. Sign in to Orris to start your own chat.