Vision: Complete Anatomy, Physiology, and Neural Pathways
Primary sources: Ganong's Review of Medical Physiology, 26th ed.; Medical Physiology (Boron & Boulpaep)
PART I - ANATOMY OF THE EYE AND OPTICS
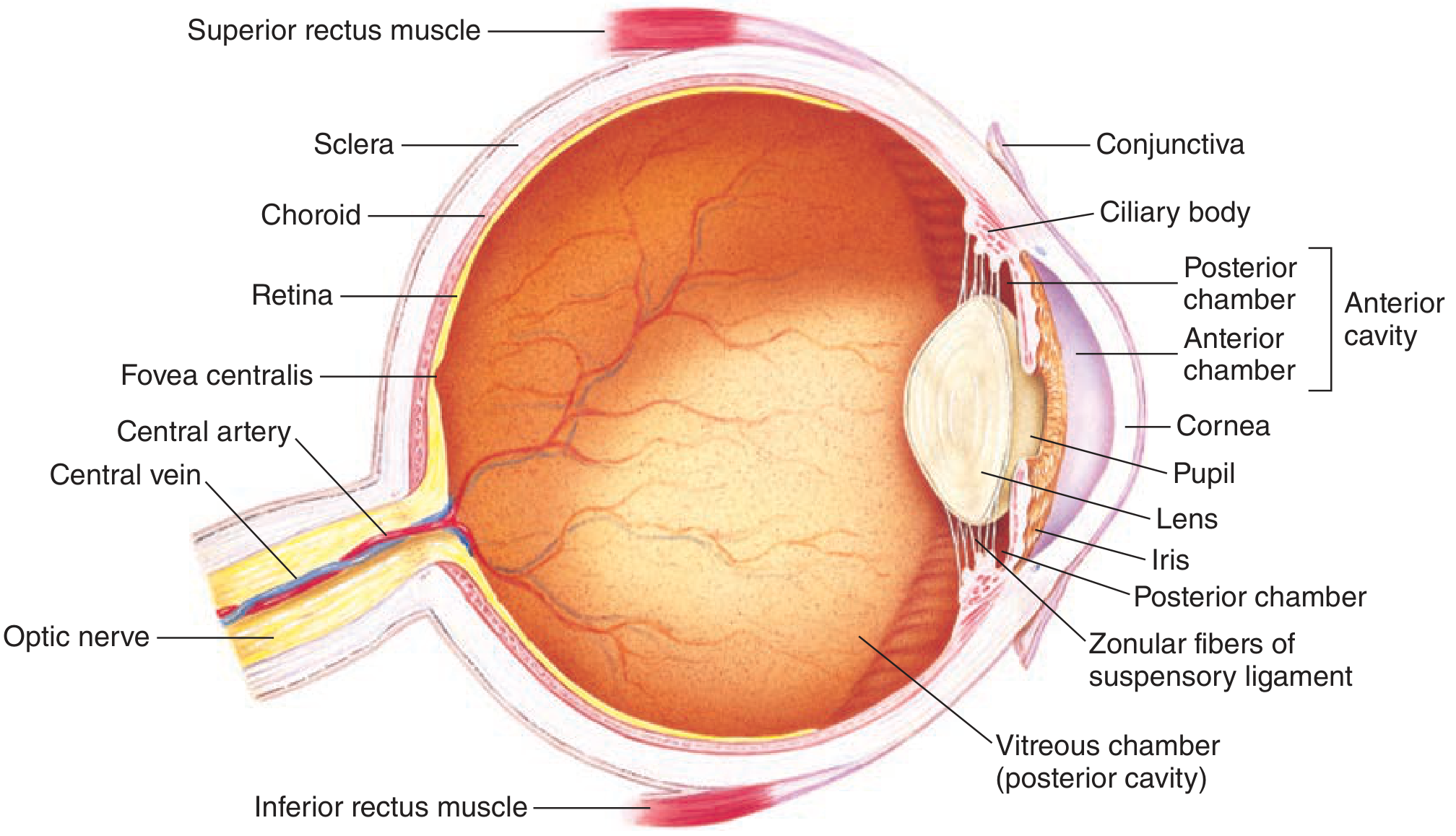
Anatomy of the Eye - Structures and Functions
Outer Coat (Fibrous Tunic)
| Structure | Function |
|---|
| Sclera | The white outer protective layer through which no light passes; maintains the shape of the eyeball and protects internal structures |
| Cornea | The transparent anterior modification of the sclera; provides ~70% of the total refractive power of the eye (~43 diopters); light rays enter here |
| Conjunctiva | Clear mucous membrane covering the sclera; contiguous with the lateral margin of the cornea; provides lubrication and protection |
Middle Coat (Uveal Tract)
| Structure | Function |
|---|
| Choroid | Highly vascular layer just inside the sclera; delivers oxygen and nutrients to the outer retina and photoreceptors |
| Ciliary body | Contains ciliary muscle (smooth muscle); controls the shape of the lens for accommodation via the zonular (suspensory) fibers |
| Iris | The colored, ring-shaped diaphragm surrounding the pupil; contains sphincter muscle (parasympathetic) for miosis and dilator muscle (sympathetic) for mydriasis; regulates light entering the eye |
| Pupil | The central aperture of the iris; adjusts diameter from ~2 mm (bright light) to ~8 mm (darkness) to regulate retinal illuminance by a factor >16x |
Inner Coat (Retina)
| Structure | Function |
|---|
| Retina | Neural tissue containing photoreceptors; lines the posterior two-thirds of the choroid; converts light to neural signals |
| Fovea centralis | Small central pit (~1.5 mm) containing only cones (no rods); responsible for highest visual acuity and color vision |
| Optic disc (blind spot) | Where ganglion cell axons exit as the optic nerve; contains no photoreceptors |
| Macula lutea | Yellow-pigmented area surrounding the fovea; region of highest cone density |
| Pigment epithelium | Absorbs light rays to prevent reflection back through the retina; phagocytoses shed rod outer segment discs; recycles retinoids |
Internal Structures
| Structure | Function |
|---|
| Lens | Biconvex transparent crystalline structure suspended by zonular fibers; provides ~30% of refractive power; changes shape during accommodation |
| Vitreous chamber (posterior cavity) | Filled with vitreous humor (gel-like); maintains the spherical shape of the eyeball and holds the retina in place |
| Aqueous humor | Fills the anterior and posterior chambers; produced by the ciliary body; maintains intraocular pressure; nourishes the avascular cornea and lens |
| Anterior chamber | Between cornea and iris; filled with aqueous humor |
| Posterior chamber | Between iris and lens; filled with aqueous humor |
| Optic nerve | Formed by ganglion cell axons (~1.2 million fibers per nerve); exits the eye at the optic disc; carries visual signals to the brain |
Extraocular Muscles
Six muscles move the eye in its orbit - superior rectus, inferior rectus, medial rectus, lateral rectus (all innervated by CN III except lateral rectus by CN VI), superior oblique (CN IV), and inferior oblique (CN III). Together they allow the four types of eye movements described below.
How Light is Brought to Focus on the Retina
The total refracting power of the eye is approximately 60 diopters. Light entering the eye is refracted (bent) at four surfaces: the anterior corneal surface, posterior corneal surface, anterior lens surface, and posterior lens surface. The cornea contributes the majority of fixed refraction (~43 D); the lens adds variable refraction (~17 D at rest, up to ~29 D during maximum accommodation).
For a relaxed eye, parallel rays from objects more than ~7 meters away converge precisely on the retina. Closer objects require greater convergence - the focal point would otherwise fall behind the retina. To keep it on the retina, the eye increases its refractive power through accommodation.
The image projected onto the retina is inverted (upside down) and reversed left-to-right relative to the external object - this is corrected by cortical processing.
Accommodation
Accommodation is the process by which the eye increases its focal power to bring nearby objects into sharp focus. It operates as follows:
- The ciliary muscle contracts (parasympathetic stimulation via CN III, ciliary ganglion)
- This releases the tension on the zonular (suspensory) fibers
- The lens, freed from radial tension, rounds up due to its inherent elasticity
- Increased curvature = increased refractive power; the focal point shifts forward to land on the retina
Young children can increase focal power by up to 12-14 diopters; their near point is a few centimeters. With age, the lens becomes stiffer and loses this elasticity, progressively reducing the range of accommodation.
The accommodation reflex involves three simultaneous components: (1) lens rounding (accommodation), (2) pupillary constriction (reduces spherical aberration), and (3) convergence of the eyes medially.
Refractive Errors
Myopia (Nearsightedness)
The anteroposterior diameter of the eyeball is too long, or the cornea is too curved. Parallel rays from distant objects focus in front of the retina, producing a blurred image. Near objects can still be seen clearly (the focal point coincidentally falls on the retina). Corrected with concave (diverging) lenses, which move the focal plane posteriorly onto the retina.
Hyperopia (Farsightedness)
The eyeball is too short, or the cornea too flat. Even with maximum lens accommodation, the focal point of near objects falls behind the retina. Young hyperopes can compensate with accommodation; older patients cannot. Corrected with convex (converging) lenses.
Astigmatism
Uneven curvature of one or more refracting surfaces (most commonly the cornea). A point source of light cannot be brought to a single precise focus - different meridians focus at different points, producing a smeared or blurred image. Corrected with cylindrical lenses that compensate for the irregular curvature.
Presbyopia
Age-related loss of accommodation caused by progressive stiffening of the lens (reduced plasticity) and weakening of the ciliary muscle. By the mid-40s, the near point exceeds arm's length. Virtually universal past middle age. Corrected with reading glasses (convex lenses) or bifocals.
Strabismus
Strabismus (squint) is a misalignment of the eyes - the visual axes do not point to the same object simultaneously. It results from imbalanced action of the extraocular muscles (due to weakness, paresis, or abnormal innervation). Types include:
- Esotropia: one eye deviates medially (convergent squint)
- Exotropia: one eye deviates laterally (divergent squint)
- Hypertropia/Hypotropia: vertical deviations
The brain receives two non-corresponding retinal images. In adults this causes diplopia (double vision). In children, the brain suppresses the image from the deviating eye - if untreated, this leads to amblyopia (lazy eye), a permanent reduction in visual acuity in the suppressed eye. Treatment includes patching the dominant eye, corrective lenses, prisms, and/or surgery on the extraocular muscles.
Neural Pathways: Rods and Cones to Visual Cortex
The pathway from photoreceptors to cortex has the following sequence:
1. Photoreceptors (Rods and Cones)
Rods and cones synapse onto bipolar cells in the outer plexiform layer of the retina.
2. Bipolar Cells → Ganglion Cells
Bipolar cells carry the signal to ganglion cells in the inner plexiform layer. The ~1.2 million ganglion cell axons converge to form the optic nerve.
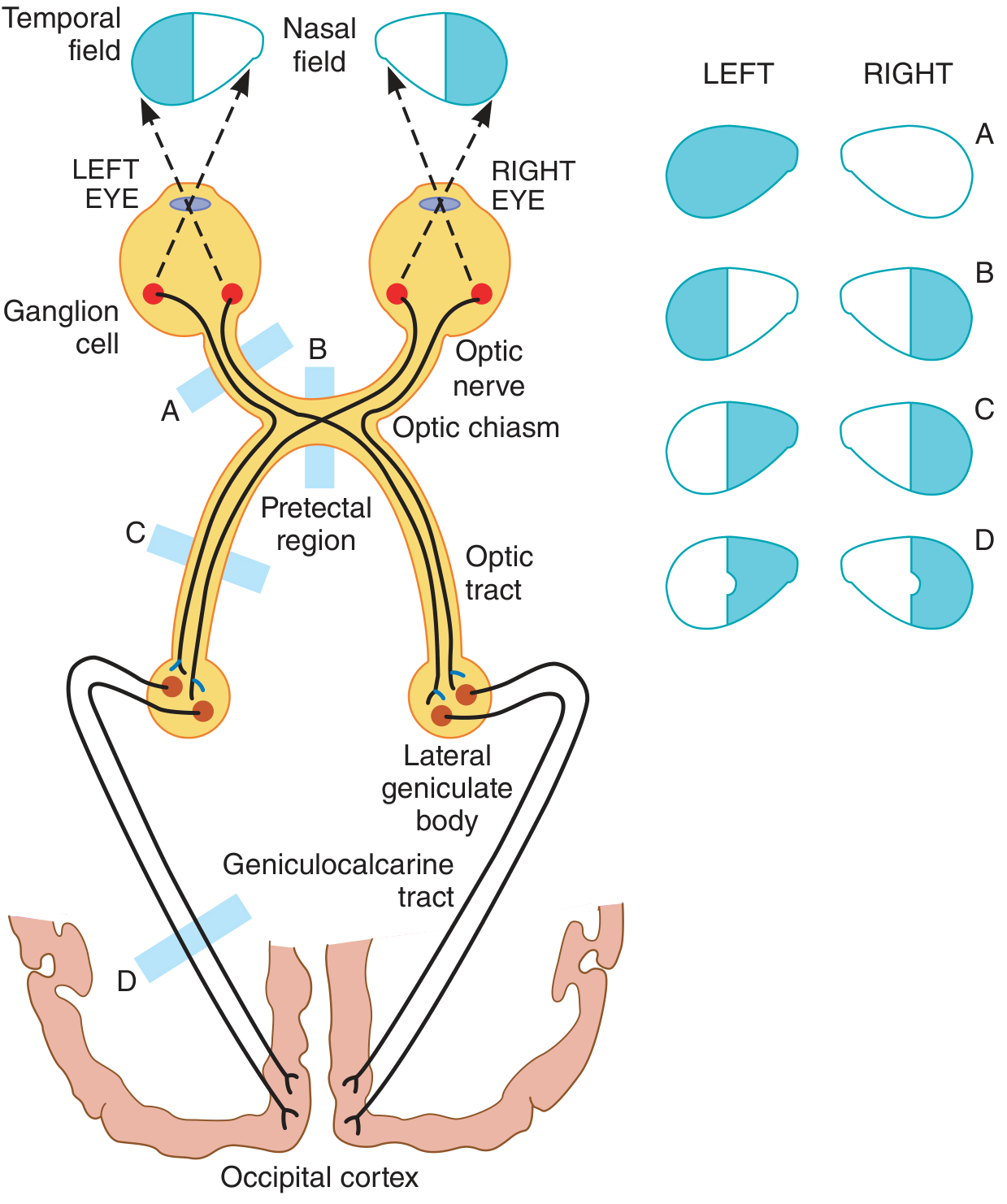
3. Optic Nerve → Optic Chiasm
At the optic chiasm, fibers from the nasal (medial) hemiretina of each eye decussate (cross) to the contralateral side. Fibers from the temporal hemiretina remain ipsilateral. This means each optic tract carries information from the contralateral visual field (temporal retina ipsilateral + nasal retina contralateral).
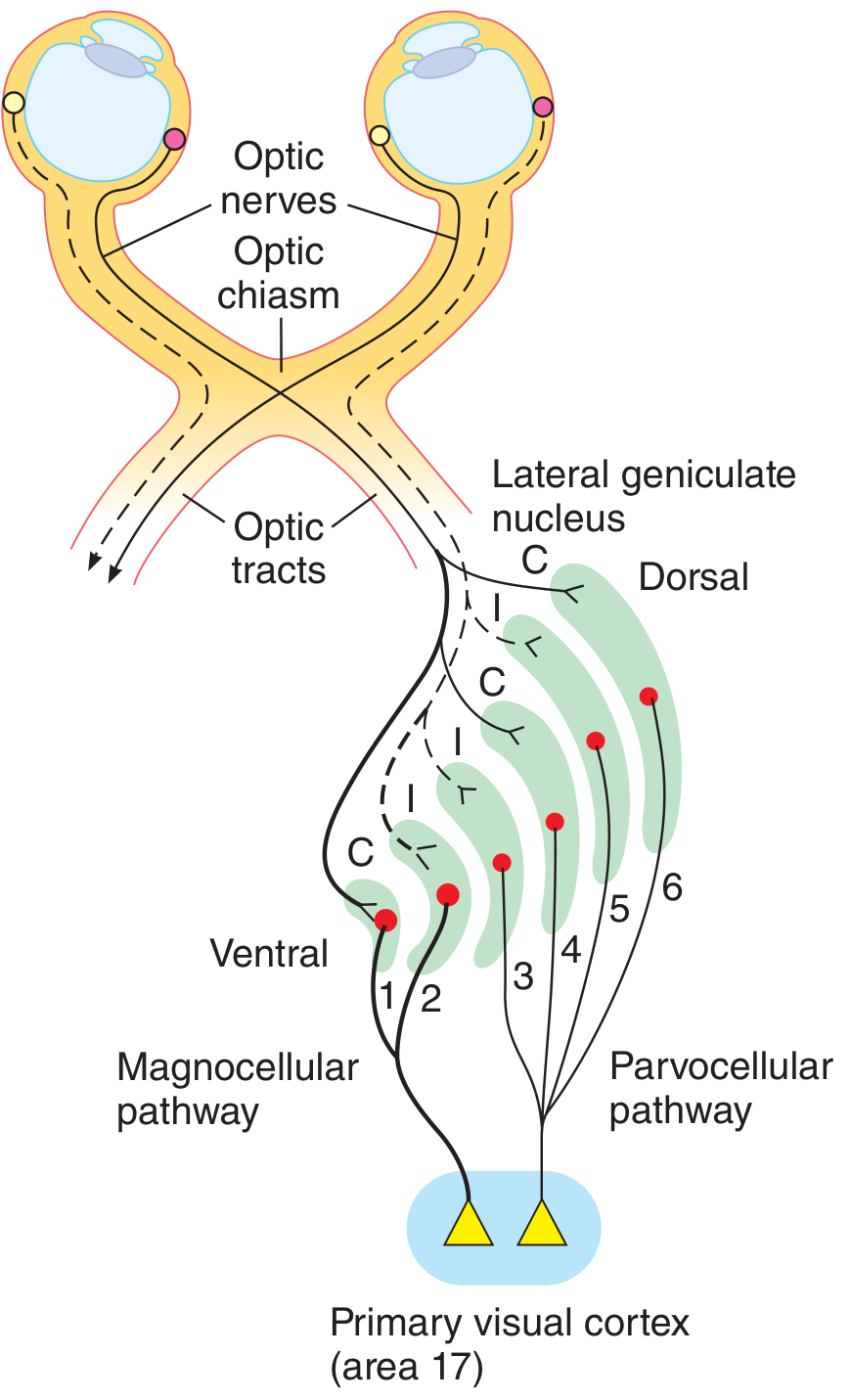
4. Optic Tract → Lateral Geniculate Body (LGB)
The optic tract terminates in the lateral geniculate nucleus (LGN) of the thalamus. The LGN has 6 well-defined layers:
- Layers 1 and 2: Magnocellular - large cells receiving input from M-type (magno) ganglion cells; process motion and spatial contrast
- Layers 3-6: Parvocellular - small cells receiving input from P-type (parvo) ganglion cells; process color, fine detail, and form
- Layers 1, 4, 6: receive input from the contralateral eye
- Layers 2, 3, 5: receive input from the ipsilateral eye
5. Geniculocalcarine Tract (Optic Radiation) → Primary Visual Cortex (V1)
LGN neurons project via the optic radiation to the primary visual cortex (V1, area 17) in the occipital lobe, along the calcarine fissure. Upper retinal quadrants project to the upper (cuneal) lip; lower quadrants project to the lower (lingual) lip. Macular (central) vision has a disproportionately large cortical representation at the occipital pole.
Lesion Effects on Visual Fields (see diagram above):
- A - Cut one optic nerve: blindness in that eye only
- B - Cut optic chiasm (e.g., pituitary tumor): bitemporal heteronymous hemianopia (loss of both temporal fields)
- C - Cut one optic tract: contralateral homonymous hemianopia
- D - Occipital lobe lesion: contralateral homonymous hemianopia, often with macular sparing
PART II - PHOTORECEPTOR AND RETINAL CELL PHYSIOLOGY
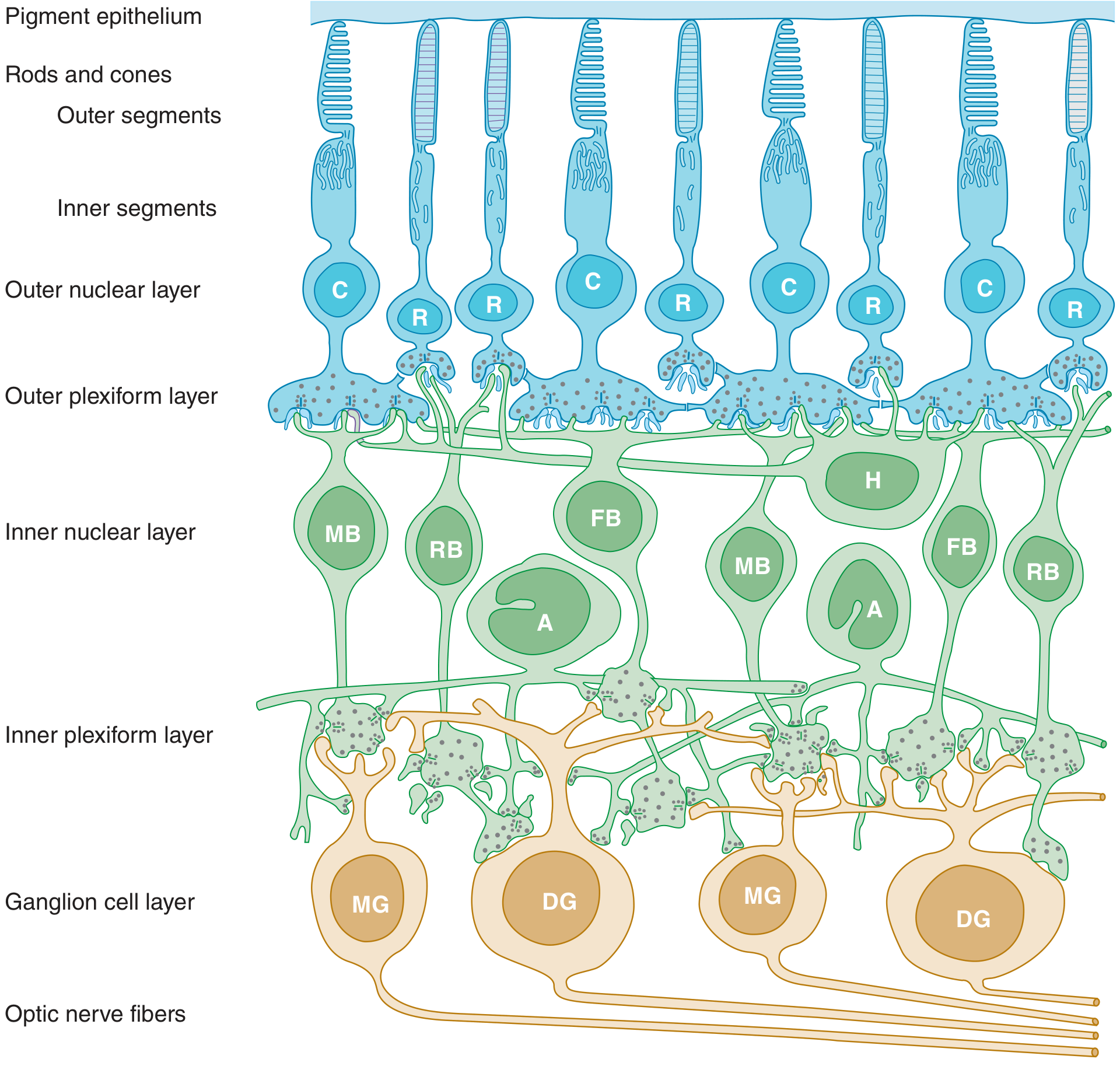
The Retinal Layers
The retina is organized into distinct layers (from outer/choroidal side inward):
- Pigment epithelium
- Photoreceptor outer and inner segments (rods = R; cones = C)
- Outer nuclear layer (photoreceptor nuclei)
- Outer plexiform layer (synapses between photoreceptors and bipolar/horizontal cells)
- Inner nuclear layer (bipolar = MB/FB/RB, horizontal = H, amacrine = A cells)
- Inner plexiform layer (synapses between bipolar/amacrine and ganglion cells)
- Ganglion cell layer (midget = MG, diffuse = DG ganglion cells)
- Optic nerve fiber layer (ganglion cell axons)
Electrical Responses of Rods and Cones (Phototransduction)
Structure
Each photoreceptor has:
- Outer segment: stacks of disc membranes (rods: free-floating discs; cones: infolded saccules) embedded with photosensitive pigment
- Inner segment: rich in mitochondria; synthesizes photopigment
- Synaptic terminal: releases glutamate
The Photosensitive Pigments
- Rods: contain rhodopsin (opsin + 11-cis retinal, a vitamin A derivative)
- Cones: contain photopsins - three types based on spectral sensitivity:
- L-cones (red-sensitive, ~564 nm peak)
- M-cones (green-sensitive, ~534 nm peak)
- S-cones (blue-sensitive, ~420 nm peak)
All cone opsins have ~41% homology with rhodopsin; the L and M opsins have ~96% homology with each other.
Phototransduction Cascade
In darkness:
- The outer segment membrane is relatively permeable to Na⁺ (and Ca²⁺) through cGMP-gated cation channels, which are held open by high intracellular cGMP levels
- This creates a steady inward "dark current"
- The photoreceptor is depolarized (membrane potential ~-40 mV)
- At the synaptic terminal, the depolarized cell continuously releases glutamate onto bipolar cells
In light:
- A photon is absorbed by the photopigment
- 11-cis retinal isomerizes to all-trans retinal, activating opsin
- Activated opsin activates transducin (a G-protein, Gₜ)
- Transducin activates phosphodiesterase (PDE)
- PDE hydrolyzes cGMP → cGMP levels fall
- cGMP-gated channels close → the dark current is interrupted
- The cell hyperpolarizes (toward -70 mV)
- Hyperpolarization reduces glutamate release at the synapse
This is unique: photoreceptors respond to their adequate stimulus (light) by hyperpolarizing, not depolarizing. The signal is graded (not all-or-nothing) and proportional to light intensity.
Recovery (dark adaptation): Rhodopsin is regenerated via phosphorylation by rhodopsin kinase and binding of arrestin, then recycling of all-trans retinal back to 11-cis retinal in the pigment epithelium.
Retinal Interneuron Responses and Functions
Bipolar Cells
Bipolar cells connect photoreceptors to ganglion cells and come in two functionally distinct types:
- ON-center bipolar cells: use mGluR6 (metabotropic) glutamate receptors. In darkness, glutamate from photoreceptors keeps them hyperpolarized. When light reduces glutamate, they depolarize (excited by light ON)
- OFF-center bipolar cells: use AMPA/kainate (ionotropic) glutamate receptors. In darkness, glutamate depolarizes them. When light reduces glutamate, they hyperpolarize (excited by light OFF)
Different subtypes include midget bipolar cells (connected to single cones in the fovea), rod bipolar cells (connected to rods), and flat (diffuse) bipolar cells (connected to multiple cones).
Horizontal Cells
- Located in the inner nuclear layer; extend laterally across the outer plexiform layer
- Receive input from photoreceptors and feed back inhibitory signals onto photoreceptors and bipolar cells
- Mediate lateral inhibition - their inhibitory feedback from illuminated surround photoreceptors onto central bipolar cells creates the center-surround receptive field of ganglion cells
- Responsible for the inhibitory surround of retinal receptive fields, which sharpens contrast and edge detection
Amacrine Cells
- Located in the inner nuclear layer; extend processes into the inner plexiform layer
- Diverse group (~30 subtypes) using various neurotransmitters (GABA, glycine, dopamine, acetylcholine, serotonin)
- Connect bipolar cells to ganglion cells; also interconnect ganglion cells with each other
- Process temporal features of the visual signal - many are sensitive to moving stimuli and changes in illumination
- Rod pathway: rod signals travel rod → rod bipolar → AII amacrine → ON/OFF cone bipolar → ganglion cell (since there is no direct rod-to-ganglion cell pathway)
- Some amacrine cells (e.g., starburst amacrine cells) are critical for direction selectivity in retinal ganglion cells
Ganglion Cells
Ganglion cells are the only output neurons of the retina; their axons form the optic nerve. They generate true action potentials. They have center-surround receptive fields (created by horizontal and amacrine cell circuitry):
- ON-center cells: excited when light hits the center of their receptive field; inhibited by light in the surround
- OFF-center cells: inhibited by central light; excited by surround light
Two major functional types:
- M-type (magno) ganglion cells (parasol cells): large cells with large receptive fields; project to LGN layers 1-2; sensitive to motion, contrast, and depth; fast-conducting; little color discrimination
- P-type (parvo) ganglion cells (midget cells): small cells with small receptive fields; project to LGN layers 3-6; sensitive to fine spatial detail and color; slow-conducting; dominate foveal output (~80% of all ganglion cells)
- Intrinsically photosensitive retinal ganglion cells (ipRGCs): contain melanopsin; project to the suprachiasmatic nucleus for circadian rhythm regulation and to the pretectal area for the pupillary light reflex
Visual Cortex Responses and Functional Organization
Primary Visual Cortex (V1, Area 17, Striate Cortex)
V1 is organized into six layers. LGN fibers terminate primarily in layer 4, which is further divided into 4A, 4B, 4Cα (magno input), and 4Cβ (parvo input).
V1 contains:
- Orientation columns: neurons respond maximally to edges/bars at a specific orientation; adjacent columns shift orientation by ~10°
- Ocular dominance columns: alternating strips of cortex preferentially responding to one eye or the other
- Blobs (cytochrome oxidase-rich patches): contain wavelength-sensitive neurons with little orientation selectivity; important for color processing
- Simple cells (respond to oriented edges at specific retinal locations), complex cells (respond to oriented edges moving in a preferred direction), and hypercomplex cells (end-stopped cells responding to edges/corners of specific lengths)
Extrastriate Visual Areas (V2-V8)
| Area | Function |
|---|
| V2, V3, VP | Continued processing; larger receptive fields |
| V3A | Motion processing |
| V4 | Detection of shape, color, and texture |
| MT/V5 | Direction of movement; motion processing |
| V6 | Direction and speed of bodily movement |
| V7 | Upper/lower quadrant visual field representation |
| V8 | Color vision |
| LO (lateral occipital) | Shape recognition |
Dorsal and Ventral Visual Pathways
From V1, visual information diverges into two major processing streams:
Dorsal Pathway ("Where/How" pathway)
- V1 → V2 → V3A → MT/V5 → posterior parietal cortex → frontal lobe
- Processes: motion, spatial location, visually guided movement, depth perception, attention
- Damage causes: optic ataxia, hemispatial neglect, simultanagnosia, geographic disorientation, dressing apraxia
Ventral Pathway ("What" pathway)
- V1 → V2 → V4 → V8 → inferior temporal cortex (IT cortex) → hippocampus/entorhinal cortex
- Processes: color, shape, form, object identity, face recognition
- Damage causes: visual agnosia (inability to recognize objects), prosopagnosia (inability to recognize faces), alexia (inability to read), achromatopsia (loss of color perception)
Color Vision Neural Pathways
Color vision begins with the three cone types (L, M, S - trichromacy). The neural processing is opponent-color:
At the ganglion cell/LGN level, three opponent pathways carry color information:
- Red-green (L-M) pathway: signals the difference between L-cone and M-cone responses. Excited by red light (L > M), inhibited by green light (M > L). Mediated primarily by P-type ganglion cells.
- Blue-yellow (S vs. L+M) pathway: signals the difference between S-cone responses and the sum of L+M responses. Excited by blue (S dominant), inhibited by yellow (L+M dominant). Mediated by small bistratified ganglion cells.
- Luminance (L+M) pathway: signals the sum of L and M cone responses; carries brightness information. This is the magnocellular pathway.
These three pathways project to blobs (cytochrome oxidase patches) and layer 4C of V1 → then to V4 and V8 for higher-order color processing. How V8 converts opponent-color input into perceived color sensations is not yet fully understood.
Color blindness results from absent or abnormal cone pigments. The most common form is red-green color blindness, caused by absence or mutation of L-cones (protanopia/protanomaly) or M-cones (deuteranopia/deuteranomaly), both X-linked recessive. S-cone (blue) defects (tritanopia) are rare and autosomal.
FOUR TYPES OF EYE MOVEMENTS
All four movement types share the same final common pathway - the motor neurons innervating the six extraocular muscles (CN III, IV, VI). They are controlled by different central neural systems.
1. Saccades
Definition: Rapid, conjugate, jerky shifts of gaze from one point to another (up to 700°/sec).
Function:
- Quickly bring a new object of interest onto the fovea
- Prevent visual adaptation (neural fatigue) that would occur if gaze were fixed on one object continuously
- Occur constantly during reading, scanning a scene, and rapid gaze shifts
Neural control:
- The frontal eye fields (FEF, area 8) and the supplementary eye fields initiate voluntary saccades
- The superior colliculus encodes the direction and amplitude of saccades
- The paramedian pontine reticular formation (PPRF) is the horizontal gaze center; the rostral interstitial nucleus of the MLF (riMLF) controls vertical saccades
- The pathway is: cortex → superior colliculus → PPRF → abducens nucleus → CN VI + contralateral CN III via the medial longitudinal fasciculus (MLF)
2. Smooth Pursuit Movements
Definition: Slow, sustained tracking movements of the eyes that follow a moving target (typically up to 30-100°/sec).
Function:
- Keep a moving object continuously imaged on the fovea
- Allow sustained focused vision of moving targets (e.g., a ball in flight, a car)
- Require a moving target as a stimulus (cannot be made voluntarily without a moving target)
Neural control:
- Require visual input from the motion-sensitive areas MT/V5
- The middle temporal (MT) and medial superior temporal (MST) areas process target velocity
- Signals pass to the frontal pursuit area and cerebellum (flocculus and vermis)
- The cerebellum is critical for smooth pursuit - cerebellar lesions produce "saccadic pursuit" (cogwheel appearance)
- Final pathway via PPRF and abducens nucleus (similar to saccades but at much lower velocity)
3. Vestibular Movements (Vestibulo-Ocular Reflex, VOR)
Definition: Compensatory eye movements that maintain a stable retinal image during head movements; driven by the vestibular system (semicircular canals and otolith organs).
Function:
- When the head rotates, the eyes move in the opposite direction with equal speed and amplitude, keeping the visual image stable on the retina
- Operates at extremely short latency (~10 ms) - faster than the visual system can respond
- Works even in darkness (no visual feedback required)
Neural control:
- Semicircular canals detect angular acceleration → signals travel via CN VIII to the vestibular nuclei
- Vestibular nuclei project directly to the oculomotor, trochlear, and abducens nuclei
- The MLF is the key pathway coordinating conjugate eye movements during head rotation
- The VOR is supplemented by the cervico-ocular reflex (neck proprioceptors) and optokinetic reflex
Clinical note: The caloric test (irrigating the ear with warm or cold water) exploits the VOR to test brainstem integrity.
4. Vergence Movements
Definition: Disjunctive eye movements - the two eyes move in opposite directions to maintain fixation on objects at different depths.
- Convergence: both eyes rotate medially (nasally) when fixating on a near object
- Divergence: both eyes rotate laterally when shifting fixation to a distant object
Function:
- Prevent diplopia (double vision) when gaze shifts between near and far objects
- Maintain corresponding retinal images (binocular fusion)
- Work together with accommodation and pupil constriction as the near triad
Neural control:
- Controlled by the mesencephalic reticular formation (near the CN III nucleus)
- The near reflex triad (accommodation + convergence + miosis) is triggered by the cortex (Area 19/V3) detecting blurring when a near object is viewed
- Signals pass through the Edinger-Westphal nucleus (for miosis and accommodation via ciliary ganglion) and medial rectus subnuclei (for convergence)
Summary Table: Eye Movements
| Movement | Speed | Binocular Pattern | Key Structure | Function |
|---|
| Saccades | Very fast (up to 700°/sec) | Conjugate (same direction) | PPRF, superior colliculus, FEF | Shift gaze; prevent adaptation |
| Smooth pursuit | Slow (up to 100°/sec) | Conjugate | MT/V5, cerebellum, PPRF | Track moving objects |
| Vestibular (VOR) | Matches head rotation | Conjugate (opposite to head) | Semicircular canals, vestibular nuclei, MLF | Stabilize gaze during head movement |
| Vergence | Slow | Disconjugate (opposite directions) | Mesencephalic RF, Edinger-Westphal nucleus | Maintain binocular fusion at different depths |
Sources: Ganong's Review of Medical Physiology, 26th ed. (McGraw-Hill) - Chapter 10; Medical Physiology, Boron & Boulpaep - Chapter 15; Eric Kandel Principles of Neural Science, 6th ed. - Chapter 21