Anatomy of inner ear in detail according to standard ent textbook, also give physiology of hearing in detail to write in pg exam of ent
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Excellent! I have all the images and content. Now let me compile the comprehensive, exam-ready answer.
Anatomy of the Inner Ear + Physiology of Hearing
(For PG ENT Exam - Based on Cummings, Scott-Brown's, KJ Lee's & Guyton)
PART 1: ANATOMY OF THE INNER EAR
Overview and Location
The inner ear (labyrinth) is housed within the petrous portion of the temporal bone. It consists of a bony labyrinth enclosing a membranous labyrinth. The bony labyrinth is filled with perilymph (similar to CSF, high Na+, low K+), and the membranous labyrinth is filled with endolymph (high K+, low Na+ - secreted by stria vascularis).
The inner ear has two functional divisions:
- Cochlea - for hearing (auditory)
- Vestibular apparatus - for balance (semicircular canals + otolith organs)
A. BONY LABYRINTH
The bony labyrinth has three parts:
1. Vestibule
- Central part of the bony labyrinth, ~4 mm in diameter
- Lies between the internal auditory meatus (anteromedially) and the middle ear cavity (laterally)
- Contains two recesses on its medial wall:
- Spherical recess (inferior/anterior) - houses the saccule
- Elliptical recess (superior/posterior) - houses the utricle
- The oval window (fenestra vestibuli) is on its lateral wall; covered by the footplate of stapes
- The cochlea sits anterior to the vestibule, connected by the ductus reuniens
- Posterior and lateral to the vestibule lie the mastoid air cells
- Medially: posterior cranial fossa, where the endolymphatic duct and sac lie beneath the dura
2. Semicircular Canals
- Three canals, each in a different plane, each opening at both ends into the vestibule
- Each canal has a dilated end - the ampulla - containing the crista ampullaris
| Canal | Plane | Notes |
|---|---|---|
| Anterior (Superior) | ~45° from sagittal plane | |
| Posterior | ~45° from sagittal plane | Shares common crus with anterior |
| Lateral (Horizontal) | Tilted 30° upward anteriorly from horizontal |
- The anterior and posterior canals share a common crus before opening into the vestibule posterosuperiorly
- The lateral canal opens independently
- Total: 5 openings from 3 canals into the vestibule
3. Cochlea
- Spiral bony canal making 2.5 to 2.75 turns around a central bony pillar called the modiolus
- Total length when uncoiled: approximately 35 mm
- Base is widest (high frequency), apex (helicotrema) is narrowest (low frequency)
- The bony spiral lamina projects from the modiolus like a screw thread, partially dividing the cochlear canal
Three compartments within the cochlea (scalae):
| Compartment | Fluid | Boundaries |
|---|---|---|
| Scala vestibuli | Perilymph | Above Reissner's membrane |
| Scala media (cochlear duct) | Endolymph | Between Reissner's & basilar membrane |
| Scala tympani | Perilymph | Below basilar membrane |
- Scala vestibuli and scala tympani communicate at the apex via the helicotrema
- Scala vestibuli opens at the oval window (covered by stapes footplate)
- Scala tympani ends at the round window (fenestra cochleae), covered by the secondary tympanic membrane
- The cochlear aqueduct (perilymphatic duct) connects scala tympani to subarachnoid space
B. MEMBRANOUS LABYRINTH
The membranous labyrinth is a closed system of fluid-filled sacs and ducts lying within the bony labyrinth. It contains endolymph.
Components:
- Cochlear duct (scala media) - in cochlea
- Utricle - in vestibule (elliptical recess)
- Saccule - in vestibule (spherical recess); connected to cochlear duct via ductus reuniens
- Three membranous semicircular ducts - within bony semicircular canals
- Endolymphatic duct and sac - extends beneath dura in posterior cranial fossa
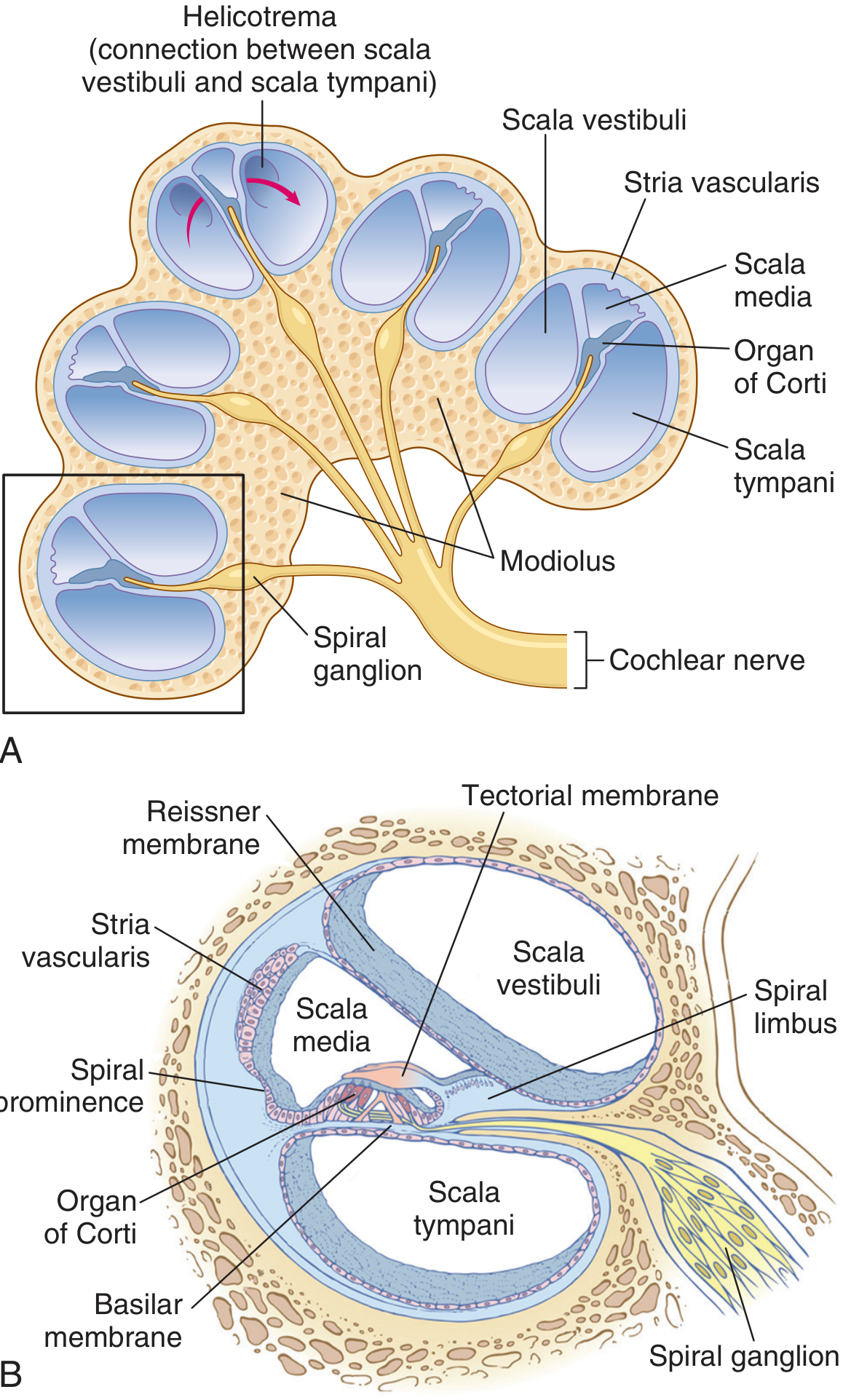
C. COCHLEAR DUCT (SCALA MEDIA) - DETAILED ANATOMY
The scala media is triangular in cross-section, bounded by:
- Roof: Reissner's membrane (vestibular membrane) - 2 layers of epithelium; so thin it doesn't impede sound transmission; maintains ionic composition of endolymph
- Outer wall: Spiral ligament + Stria vascularis (the metabolic engine - secretes endolymph and maintains +80 mV endocochlear potential)
- Floor: Basilar membrane (bearing the organ of Corti) + osseous spiral lamina
D. BASILAR MEMBRANE
- Extends from the spiral lamina (modiolus) to the spiral ligament on the outer wall
- Contains 20,000-30,000 basilar fibers
- Tonotopic organization - critical for frequency analysis:
- Base (near oval window): short, stiff, narrow fibers → responds to HIGH frequencies (20,000 Hz)
- Apex (helicotrema): long, wide, floppy fibers → responds to LOW frequencies (20 Hz)
- Stiffness decreases ~100-fold from base to apex
E. ORGAN OF CORTI
The organ of Corti sits on the basilar membrane within the scala media. It is the actual sensory transducer for hearing.
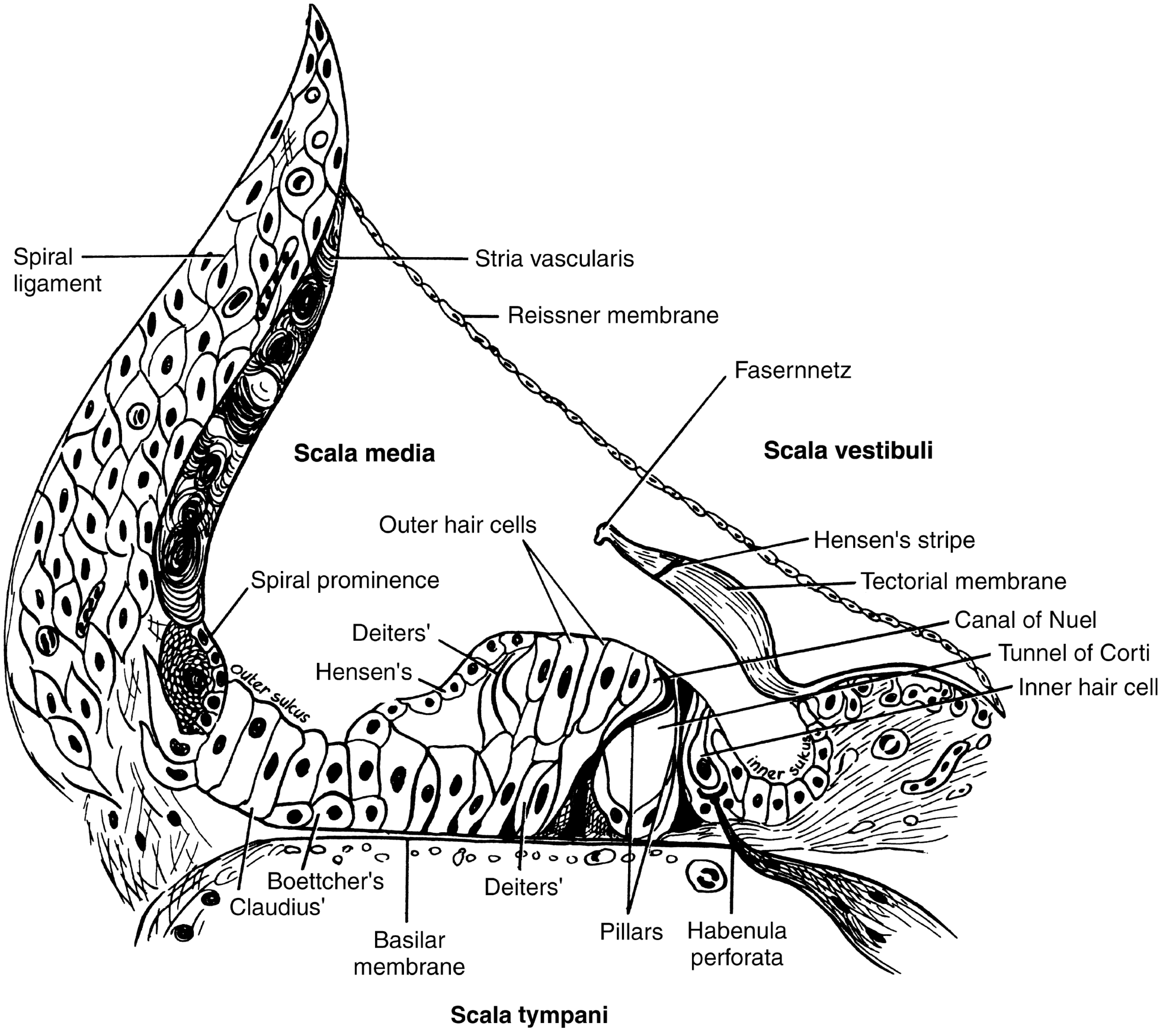
Key structures:
| Structure | Details |
|---|---|
| Inner hair cells (IHC) | Single row, ~3,500 total; each has ~50-70 stereocilia; PRIMARY afferent transducers (95% of cochlear nerve fibers) |
| Outer hair cells (OHC) | 3 rows, ~12,000-15,000 total; motile "amplifiers"; responsible for sharp frequency tuning; receive mainly efferent innervation |
| Tunnel of Corti | Space between inner and outer pillar cells; filled with cortilymph |
| Canal of Nuel | Space between outer pillar cells and outer hair cells |
| Tectorial membrane | Gelatinous membrane attached to spiral limbus; overlies and makes contact with stereocilia of OHC (and perhaps IHC); deflects stereocilia during basilar membrane movement |
| Reticular lamina | Tight junctions sealing the top of hair cells; separates endolymph (above) from perilymph (below) |
| Inner/Outer pillar cells | Form the tunnel of Corti; provide structural support |
| Deiters' cells | Support OHCs; cup-shaped phalangeal cells |
| Hensen's cells | Lateral to Deiters' cells |
| Claudius' cells | Further lateral |
| Boettcher's cells | At the base of Claudius' cells |
| Habenula perforata | Perforations in the osseous spiral lamina through which cochlear nerve fibers pass to IHCs |
F. VESTIBULAR END ORGANS
1. Semicircular Canals - Crista Ampullaris
- Located in the ampulla of each semicircular canal
- Contains hair cells with stereocilia + one kinocilium embedded in cupula (gelatinous mass)
- The cupula has the same specific gravity as endolymph - responds to angular acceleration
- Deflection of cupula toward utricle (utriculopetal) = excitation in lateral canal
- Deflection away from utricle (utriculofugal) = excitation in vertical canals
2. Utricle and Saccule - Macula
- Utricular macula: roughly horizontal plane - detects horizontal linear acceleration and head tilt
- Saccular macula: roughly vertical plane - detects vertical linear acceleration and gravity
- Hair cells embedded in gelatinous otolithic membrane containing otoconia (calcium carbonate crystals)
- The striola divides each macula; hair cell polarity reverses across the striola
G. FLUIDS OF THE INNER EAR
| Property | Perilymph | Endolymph |
|---|---|---|
| Location | Scala vestibuli, scala tympani, bony labyrinth | Scala media, membranous labyrinth |
| Na+ | High (~140 mEq/L) | Low (~1 mEq/L) |
| K+ | Low (~5 mEq/L) | High (~150 mEq/L) |
| Protein | High | Low |
| Origin | Plasma ultrafiltrate + CSF via cochlear aqueduct | Secreted by stria vascularis |
| Electrical potential | 0 mV (reference) | +80 mV (endocochlear potential) |
H. INNERVATION
-
Cochlear nerve (lower division of CN VIII): cell bodies in spiral (cochlear) ganglion within the modiolus; ~30,000 bipolar neurons
- Type I neurons (90-95%): large, myelinated; supply IHCs (monosynaptic)
- Type II neurons (5-10%): small, unmyelinated; supply OHCs
-
Vestibular nerve (upper division of CN VIII): cell bodies in Scarpa's (vestibular) ganglion in the internal auditory canal
- Superior vestibular ganglion: innervates utricle, anterior/lateral SCC cristae
- Inferior vestibular ganglion: innervates saccule, posterior SCC crista
-
Efferent innervation (Olivocochlear bundle of Rasmussen):
- Lateral olivocochlear (LOC): to IHC afferent dendrites
- Medial olivocochlear (MOC): directly to OHC soma; modulates OHC motility (feedback system)
I. BLOOD SUPPLY
- Labyrinthine artery (internal auditory artery): branch of anterior inferior cerebellar artery (AICA) in 83%, or basilar artery
- Divides into:
- Cochlear artery → spiral modiolar artery → radiating arterioles → stria vascularis
- Anterior vestibular artery → utricle, anterior/lateral SCC
- Posterior vestibular artery → saccule, posterior SCC
- No autoregulation - highly susceptible to ischemia; terminal circulation (no anastomoses)
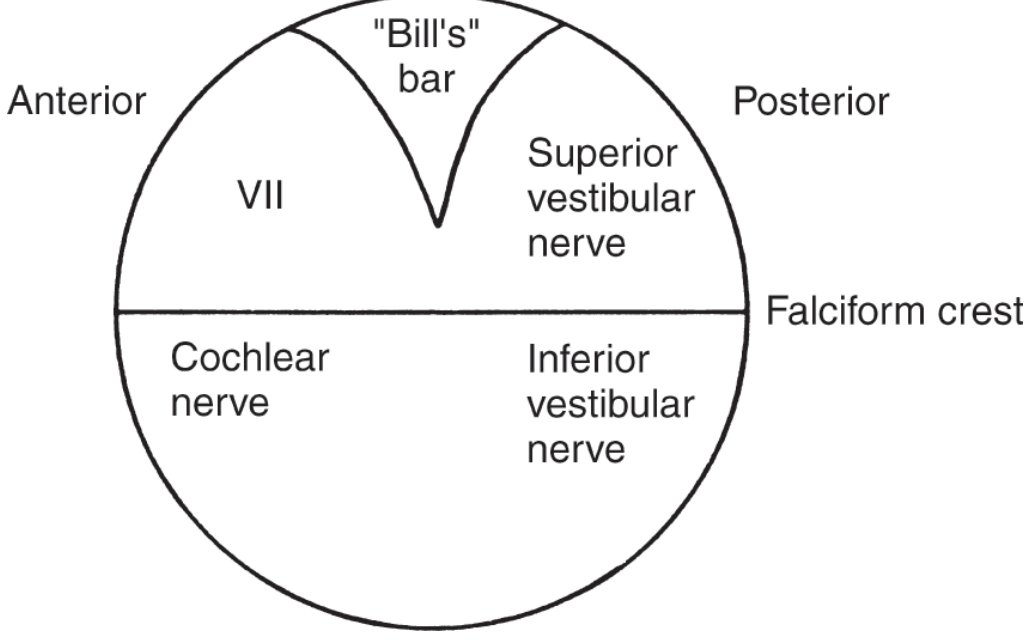
J. INTERNAL AUDITORY CANAL (IAC)
- Approximately 8-10 mm long
- Contains (Bill's bar divides superior from inferior compartment):
- Superior anterior: facial nerve (VII)
- Superior posterior: superior vestibular nerve
- Inferior anterior: cochlear nerve
- Inferior posterior: inferior vestibular nerve (singular nerve to posterior SCC)
PART 2: PHYSIOLOGY OF HEARING
Overview - The Pathway of Sound
Sound waves → External ear → Middle ear (mechanical amplification) → Oval window → Cochlear fluid waves → Basilar membrane vibration → Hair cell mechanoelectrical transduction → Cochlear nerve → Brainstem → Auditory cortex
A. IMPEDANCE MATCHING BY THE MIDDLE EAR
Sound travels through air (low impedance) to cochlear fluids (high impedance). Without impedance matching, 99.9% of sound energy (30 dB loss) would be reflected. The middle ear overcomes this by:
- Tympanic membrane to stapes footplate area ratio: Effective vibrating area of TM ~55 mm²; stapes footplate ~3.2 mm²; ratio = ~17:1 → pressure amplification of 17x
- Ossicular lever action: malleus handle: incus long process ratio = ~1.3:1
- Combined amplification: ~17 × 1.3 = 22-fold pressure gain ≈ 27 dB
This exactly compensates for the air-fluid impedance mismatch (~28 dB). The middle ear is thus a transformer or impedance-matching device.
B. COCHLEAR HYDRODYNAMICS - TRAVELING WAVE THEORY (von Bekesy, 1960 Nobel Prize)
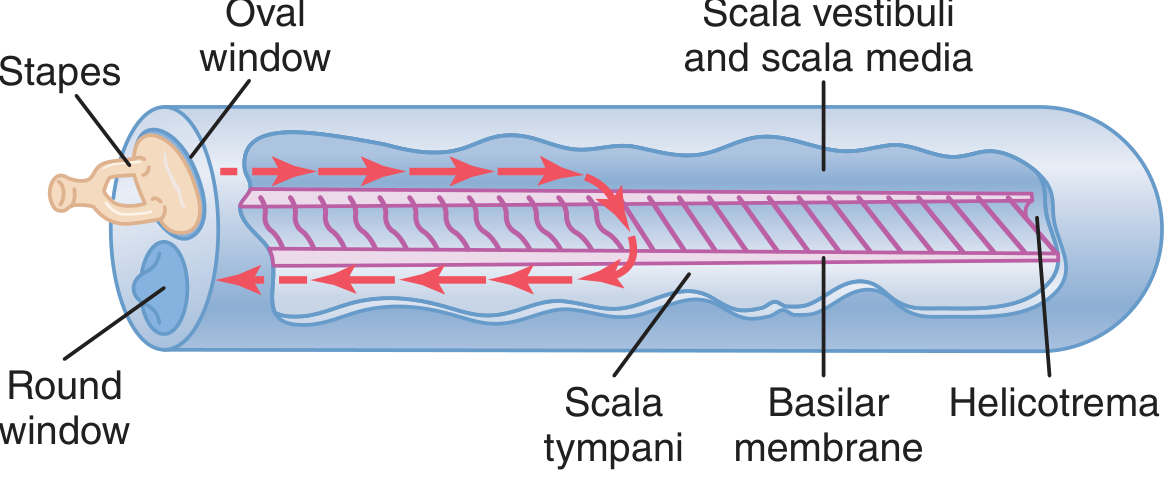
Mechanism:
- Inward movement of stapes footplate at oval window → fluid displacement in scala vestibuli
- Reissner's membrane is so thin it does not obstruct; scala vestibuli + scala media act as one chamber
- Pressure wave travels through cochlear fluid → causes traveling wave along the basilar membrane
- The wave travels from base to apex, building up in amplitude to a maximal point, then rapidly dying away
- The point of maximal displacement depends on sound frequency (Place Principle)
C. PLACE PRINCIPLE (Tonotopy) - Frequency Coding
- High frequencies: maximal vibration at the base of basilar membrane (short, stiff fibers)
- Low frequencies: maximal vibration at the apex (long, floppy fibers)
- 20 Hz → apex; 20,000 Hz → base
- The basilar membrane acts as a mechanical frequency analyzer
- This tonotopic map is preserved all the way to the auditory cortex
D. ACTIVE AMPLIFICATION - COCHLEAR AMPLIFIER (OHC Motility)
Von Bekesy's passive traveling wave theory could not explain the sharp frequency tuning or the extreme sensitivity of the cochlea. The missing element is active amplification:
- Outer hair cells (OHC) contain prestin (motor protein in the lateral wall) - allows OHCs to change length in response to receptor potentials (electromotility)
- When the basilar membrane moves, OHCs amplify this movement by up to 100-fold (40 dB gain)
- This active process sharpens the frequency tuning curve, making the cochlea extremely frequency-selective
- OHC motility is regulated by the medial olivocochlear efferent system (feedback from brainstem)
- Otoacoustic emissions (OAEs) - sounds emitted by the cochlea - are a direct consequence of OHC motility and are used clinically to test OHC function (newborn hearing screening)
E. MECHANOELECTRICAL TRANSDUCTION AT THE HAIR CELL
Step-by-step transduction:
- Basilar membrane moves upward (toward scala vestibuli)
- This causes shearing motion between the basilar membrane and the overhanging tectorial membrane
- Stereocilia of hair cells are deflected in the excitatory direction (toward the tallest stereocilium/kinocilium)
- Tip links (extracellular filaments connecting adjacent stereocilia tips) are stretched
- Mechanical stretching directly opens mechanically gated K+ channels (MET channels) at the tips of shorter stereocilia
- K+ and Ca²+ flow into the hair cell from the endolymph (high K+ in endolymph, negative intracellular)
- Hair cell depolarizes
- Depolarization opens voltage-gated Ca²+ channels at the basolateral membrane
- Ca²+ triggers exocytosis of neurotransmitter (glutamate) at the ribbon synapses
- Glutamate activates AMPA receptors on cochlear nerve afferents → action potentials → cochlear nerve
Repolarization:
- K+ exits through basolateral K+ channels into perilymph
- K+ is recycled back to endolymph via supporting cells and stria vascularis (ionic recycling via gap junctions - connexin 26 and 30)
F. ENDOCOCHLEAR POTENTIAL (ECP)
- The stria vascularis maintains a +80 mV electrical potential in the endolymph (endocochlear potential, ECP)
- The intracellular potential of hair cells is -70 mV relative to perilymph
- Therefore, across the apical membrane (stereocilia tips in endolymph), the total potential = -70 - (+80) = -150 mV
- This enormous 150 mV driving force is what makes the cochlea so exquisitely sensitive - it maximizes K+ flow through MET channels with minimal mechanical force
The ECP is generated by:
- K+ secretion by the stria vascularis (requires Na-K-ATPase)
- A highly vascular, metabolically active lateral wall structure
G. FREQUENCY PRINCIPLE (VOLLEY PRINCIPLE) - Low Frequency Coding
For very low frequencies (<200 Hz) that stimulate the same broad apical region, frequency discrimination uses:
- Phase-locking: individual nerve fibers fire in synchrony with the sound wave
- Volley principle: groups of fibers together fire volleys of impulses at the stimulus frequency
- Applicable for frequencies 20-1500 Hz
- The cochlear nucleus decodes these temporal patterns
H. LOUDNESS CODING
Loudness is encoded by three mechanisms:
- Rate coding: louder sounds cause higher firing rates of individual nerve fibers (more receptor potentials per unit time)
- Population coding: louder sounds vibrate a wider area of basilar membrane, recruiting more nerve fibers
- Threshold differences: high-threshold, high-spontaneous-rate fibers are activated only at high intensities
I. AUDITORY NEURAL PATHWAY
Organ of Corti
↓
Spiral ganglion (cochlear nerve - CN VIII)
↓
Cochlear nuclei (dorsal & ventral) - PONTOMEDULLARY JUNCTION
↓ (bilateral projections)
Superior olivary complex (SOC) - first binaural integration; localization of sound; gives off olivocochlear efferents
↓
Lateral lemniscus → Nucleus of lateral lemniscus
↓
Inferior colliculus (MIDBRAIN) - integration center; acoustic reflexes
↓
Medial geniculate nucleus (MGN) of THALAMUS
↓
Primary auditory cortex (A1) - Heschl's gyrus, superior temporal gyrus (Brodmann areas 41, 42)
Key points for exam:
- First crossing occurs at the trapezoid body (within brainstem)
- Both ipsilateral and contralateral projections exist from cochlear nuclei onwards
- Unilateral cortical lesion does NOT cause complete deafness (bilateral representation)
- Bilateral cortical lesions → cortical deafness
- Wernicke's area (area 22) - speech comprehension
J. ACOUSTIC REFLEXES
1. Stapedius Reflex (Acoustic Reflex)
- Afferent: cochlear nerve → cochlear nucleus
- Efferent: facial nerve (VII) → stapedius muscle
- Threshold: 70-90 dB above hearing threshold
- Protective - reduces ossicular chain mobility for loud sounds
- Ipsilateral and contralateral pathways
- Clinical use: tympanometry, localizing facial nerve lesions
2. Tensor Tympani Reflex
- Less reliable in humans; trigeminal nerve (V3)
- Responds to sudden non-auditory stimuli (tactile)
K. OTOACOUSTIC EMISSIONS (OAEs)
A direct byproduct of OHC active amplification:
- Spontaneous OAEs: in absence of stimulus; present in 50-70% of normal ears
- Transient evoked OAEs (TEOAEs): in response to click stimuli; used in newborn hearing screening
- Distortion product OAEs (DPOAEs): two pure tones generate distortion products; highly frequency-specific
- OAEs are absent when OHCs are damaged (noise trauma, ototoxicity); present when conductive loss is the issue
QUICK REVISION SUMMARY TABLE
| Aspect | Key Fact |
|---|---|
| Cochlear turns | 2.5-2.75 turns |
| Total cochlear length (uncoiled) | ~35 mm |
| Basilar membrane fibers | 20,000-30,000 |
| Base of BM | Short, stiff → HIGH frequency (20,000 Hz) |
| Apex of BM | Long, floppy → LOW frequency (20 Hz) |
| IHC number | ~3,500; single row; primary afferent |
| OHC number | ~12,000-15,000; 3 rows; active amplifier |
| Endocochlear potential | +80 mV (generated by stria vascularis) |
| Hair cell transduction | K+ enters through MET channels; Glutamate released |
| Neurotransmitter | Glutamate (at hair cell-afferent synapse) |
| Frequency coding | Place principle (high freq base, low freq apex) + Volley principle |
| OHC motor protein | Prestin |
| Olivocochlear bundle | Efferent feedback; modulates OHC; Rasmussen's bundle |
| Stapedius reflex arc | CN VIII → cochlear nucleus → CN VII → stapedius |
| Nobel Prize | Georg von Bekesy, 1960 - traveling wave theory |
Sources:
- Cummings Otolaryngology Head and Neck Surgery (Chapter 129 - Vestibular System; Chapter 135 - Imaging)
- Scott-Brown's Otorhinolaryngology Head & Neck Surgery, Vol 2 - Applied Physiology of Hearing
- KJ Lee's Essential Otolaryngology, 11th Ed
- Guyton & Hall Textbook of Medical Physiology (Chapter 53 - The Sense of Hearing)
This is a shared conversation. Sign in to Orris to start your own chat.