Fat and lipids chemistry and metabolism
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
Finding Sources
Loading Image
Loading Image
Loading Image
Loading Image
Reading File
I now have comprehensive content from the textbooks. Let me compile the full, structured response on fat and lipid chemistry and metabolism.
Fat and Lipids: Chemistry and Metabolism
Based on Lippincott's Illustrated Reviews in Biochemistry (8th ed.), Ganong's Review of Medical Physiology (26th ed.), and Guyton & Hall Textbook of Medical Physiology.
1. Classification and Structure of Lipids
Lipids are hydrophobic or amphipathic organic molecules. Their hydrophobic nature makes them ideal for energy storage, membrane formation, and signaling. The major classes are:
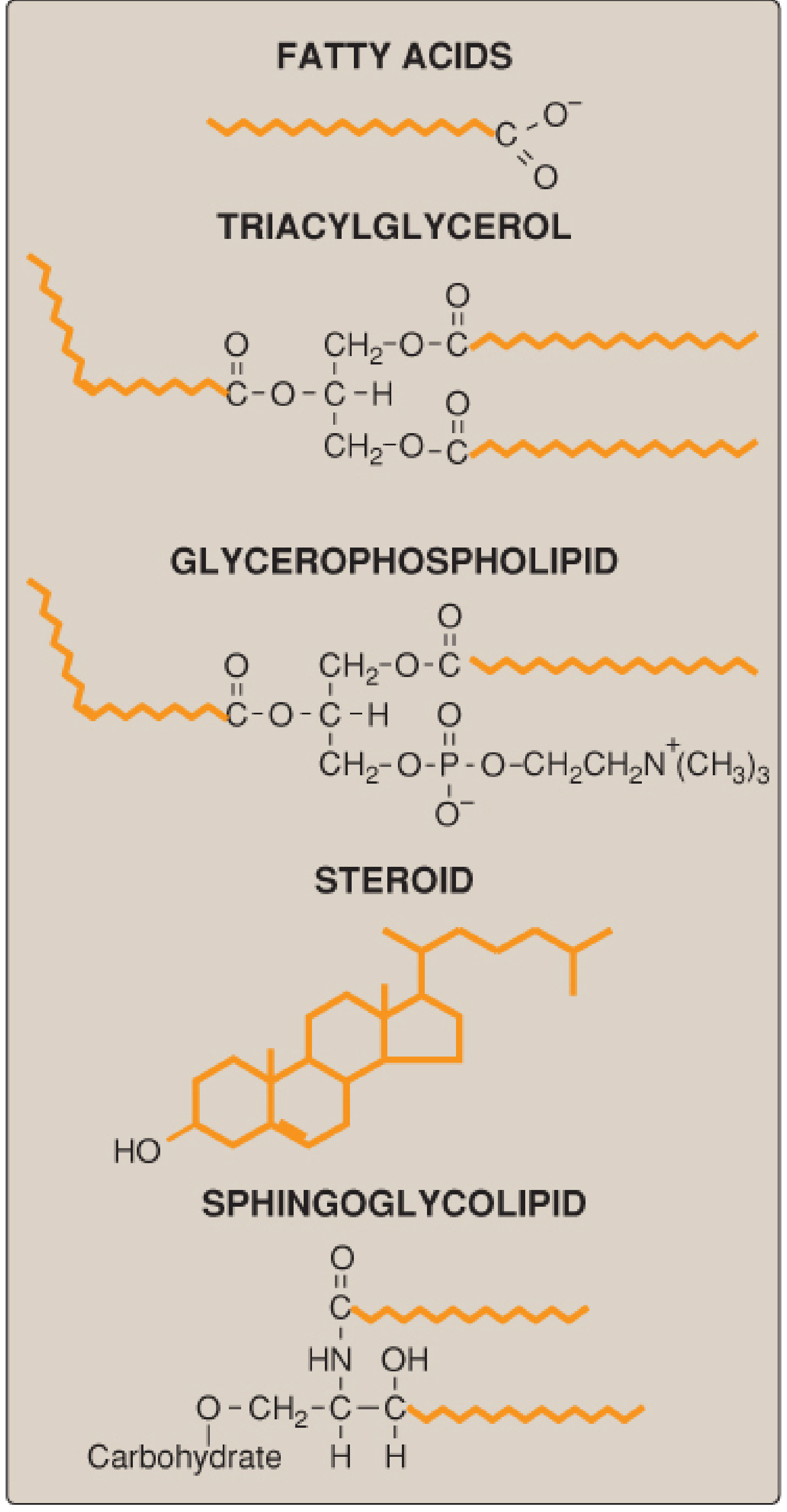
Figure: Structures of some common classes of lipids. Hydrophobic portions are shown in orange. - Lippincott Biochemistry, 8e
| Class | Core Structure | Example |
|---|---|---|
| Fatty acids | Long hydrocarbon chain + carboxyl group | Palmitic acid (16:0), Oleic acid (18:1) |
| Triacylglycerols (TAG) | Glycerol + 3 esterified fatty acids | Dietary fat; major energy store |
| Glycerophospholipids | Glycerol + 2 FA + phosphate + polar head | Phosphatidylcholine (lecithin) |
| Sphingolipids | Sphingosine backbone | Sphingomyelin, gangliosides |
| Sterols | 4-ring steroid nucleus + OH at C3 | Cholesterol |
| Waxes | Long-chain FA + long-chain alcohol | Cerumen, skin surface lipids |
Fatty Acid Nomenclature
Fatty acids are designated as C:D(Δn) where C = number of carbons, D = double bonds, n = position of first double bond from the carboxyl end.
- Saturated FA: No double bonds (e.g., palmitate 16:0, stearate 18:0)
- Monounsaturated FA: One double bond (e.g., oleic acid 18:1Δ9)
- Polyunsaturated FA (PUFA): Multiple double bonds. Two essential PUFAs (cannot be synthesized de novo in humans):
- Linoleic acid (18:2, ω-6) - precursor of arachidonic acid (20:4, ω-6), a precursor of prostaglandins/leukotrienes
- α-Linolenic acid (18:3, ω-3)
ω (omega) notation: The ω-6 designation means the first double bond from the methyl (ω) end is at carbon 6. Arachidonic acid (20:4[5,8,11,14]) is an ω-6 fatty acid because the first double bond from the ω end is 6 carbons in. - Lippincott Biochemistry, 8e
2. Lipid Digestion and Absorption
The average U.S. adult consumes ~78 g of lipid daily, >90% as TAG. Digestion begins in the stomach and is completed in the small intestine.
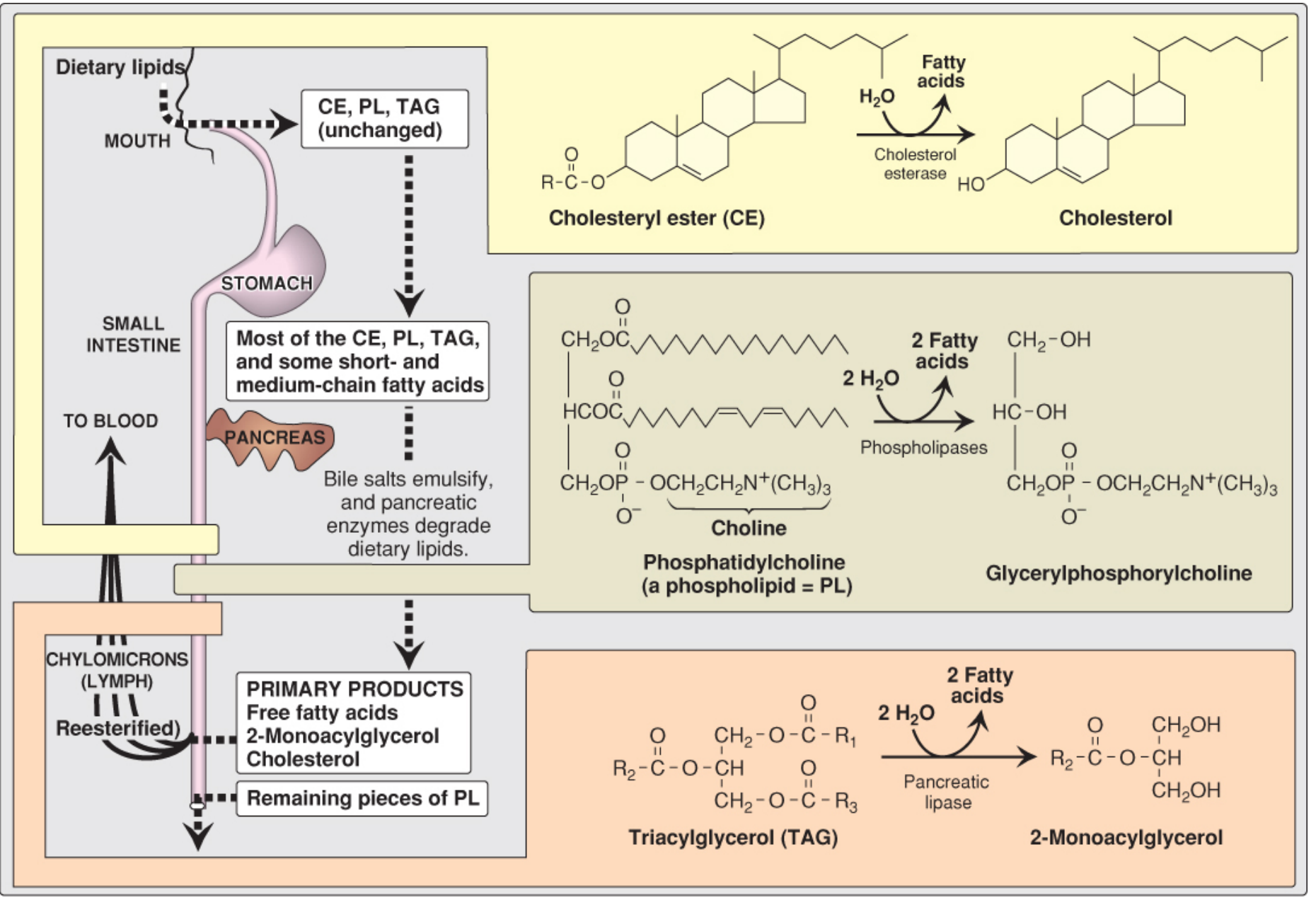
Figure: Overview of lipid digestion - Lippincott Biochemistry, 8e
Key Steps
-
Stomach: Lingual and gastric lipases (pH optimum 4-6) hydrolyze short/medium-chain FA from TAG. These enzymes are important in infants and in pancreatic insufficiency (e.g., cystic fibrosis).
-
Emulsification in the small intestine: Bile salts from the gallbladder act as detergents, breaking large fat globules into small micelles, vastly increasing the surface area for pancreatic enzymes. Cholecystokinin (CCK) triggers gallbladder contraction and release of pancreatic enzymes.
-
Enzymatic digestion:
- Pancreatic lipase (assisted by colipase) cleaves TAG at positions sn-1 and sn-3, yielding 2-monoacylglycerol + 2 free fatty acids
- Cholesterol esterase hydrolyzes cholesteryl esters to free cholesterol
- Phospholipases cleave phospholipids to lysophospholipids + FA
-
Absorption into enterocytes: Products are taken up as mixed micelles. Inside the enterocyte:
- TAG is resynthesized from 2-monoacylglycerol and FA
- Short/medium-chain FA (≤12C) pass directly into the portal blood
- Long-chain FA are repackaged into chylomicrons
-
Chylomicron secretion: Chylomicrons enter intestinal lymphatics (lacteals) and drain via the thoracic duct into the subclavian vein.
3. Lipoprotein Transport
Because lipids are hydrophobic, they are transported in the blood as lipoproteins - spherical particles with a hydrophobic core (TAG, cholesteryl esters) surrounded by a monolayer of phospholipids, cholesterol, and apolipoproteins.
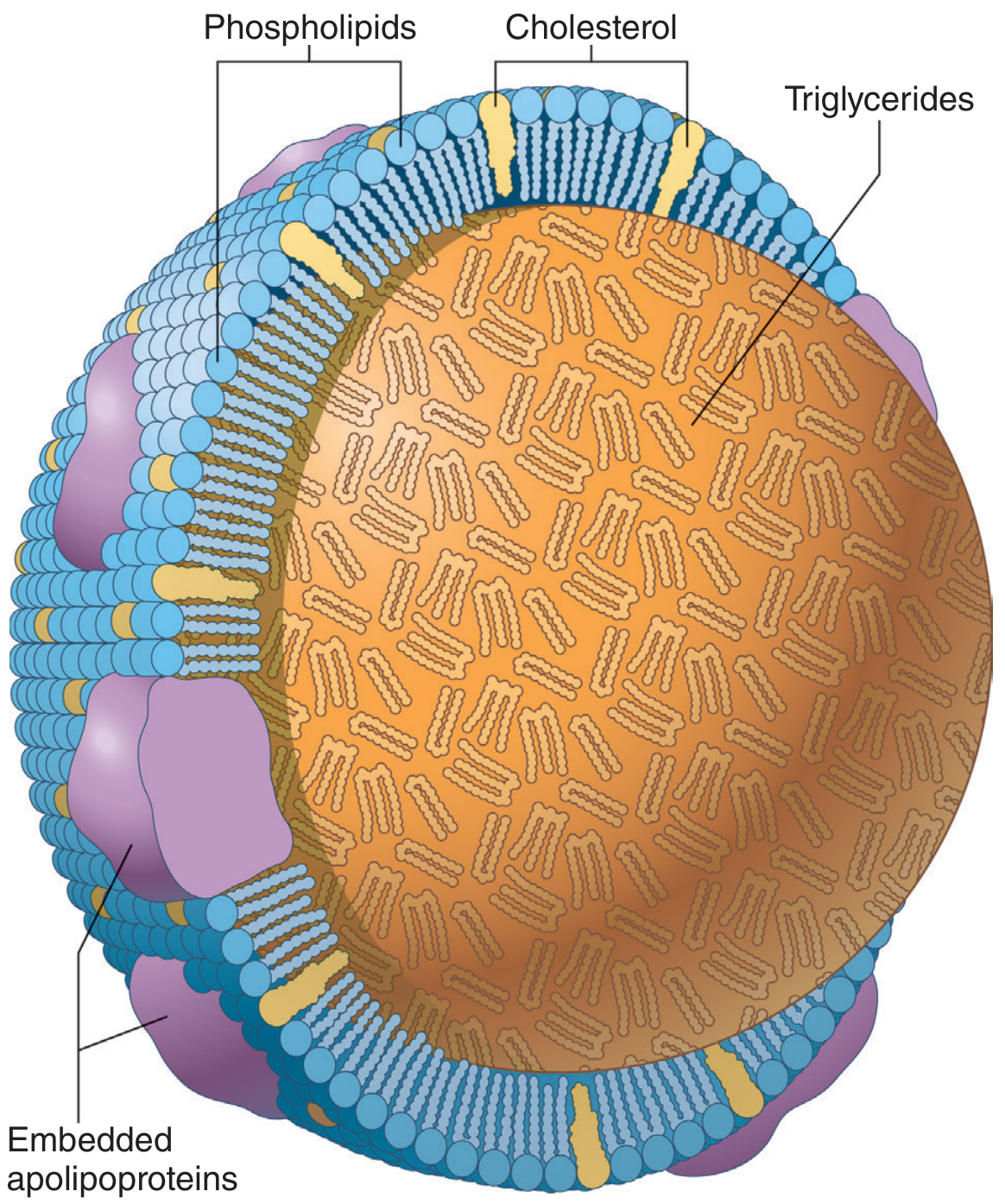
Figure: Chylomicron structure - Guyton & Hall Medical Physiology
Lipoprotein Classes (decreasing size / increasing density)
| Lipoprotein | Major Lipid | Key Apolipoproteins | Function |
|---|---|---|---|
| Chylomicron | TAG (dietary) | ApoB-48, ApoC-II, ApoE | Transport dietary fat from gut to tissues |
| VLDL | TAG (hepatic) | ApoB-100, ApoC-II, ApoE | Transport endogenous TAG from liver |
| IDL | TAG + CE | ApoB-100, ApoE | Chylomicron/VLDL remnant |
| LDL | Cholesteryl ester | ApoB-100 | Delivers cholesterol to peripheral tissues; atherogenic |
| HDL | Phospholipids/CE | ApoA-I | Reverse cholesterol transport; atheroprotective |
Chylomicron Metabolism
- After a fatty meal, chylomicrons rise to 1-2% of total plasma and plasma turns turbid/yellow.
- Chylomicron half-life is <1 hour - plasma clears within a few hours.
- Lipoprotein lipase (LPL) on capillary endothelium of adipose tissue, skeletal muscle, and heart hydrolyzes TAG in chylomicrons, releasing FA and glycerol. ApoC-II is the obligatory activator of LPL.
- FA enter adipocytes (storage) or muscle (oxidation); glycerol goes to the liver.
- Chylomicron remnants (depleted of TAG, enriched in cholesteryl esters) are taken up by hepatic ApoE receptors. - Guyton & Hall, Lippincott Biochemistry
VLDL to LDL Pathway
The liver packages endogenous TAG and cholesterol into VLDL (with ApoB-100). VLDL is progressively stripped of TAG by LPL, passing through IDL to form LDL. LDL (the major cholesterol carrier) is cleared from plasma by LDL receptors (LDLR) via endocytosis, principally in the liver. Statins work by inhibiting HMG-CoA reductase, upregulating LDL receptors and lowering plasma LDL.
4. Fatty Acid Oxidation (β-Oxidation)
Activation and Transport into Mitochondria
- FA are first activated to acyl-CoA by acyl-CoA synthetase (thiokinase) at the outer mitochondrial membrane, consuming 2 ATP equivalents (ATP → AMP + PPi).
- Long-chain acyl-CoA cannot cross the inner mitochondrial membrane directly. They are transferred to carnitine by carnitine acyltransferase I (CAT I) on the outer membrane - this is the rate-limiting step of FA oxidation.
- The acylcarnitine is transported across the membrane via a translocase, and the acyl group is transferred back to CoA inside the matrix by CAT II.
- Short- and medium-chain FA can enter mitochondria without carnitine.
Carnitine is β-hydroxy-γ-trimethylammonium butyrate, synthesized in the body from lysine and methionine. - Ganong's Review of Medical Physiology, 26e
The Four-Step β-Oxidation Spiral
Each cycle removes a 2-carbon acetyl-CoA unit from the acyl chain:
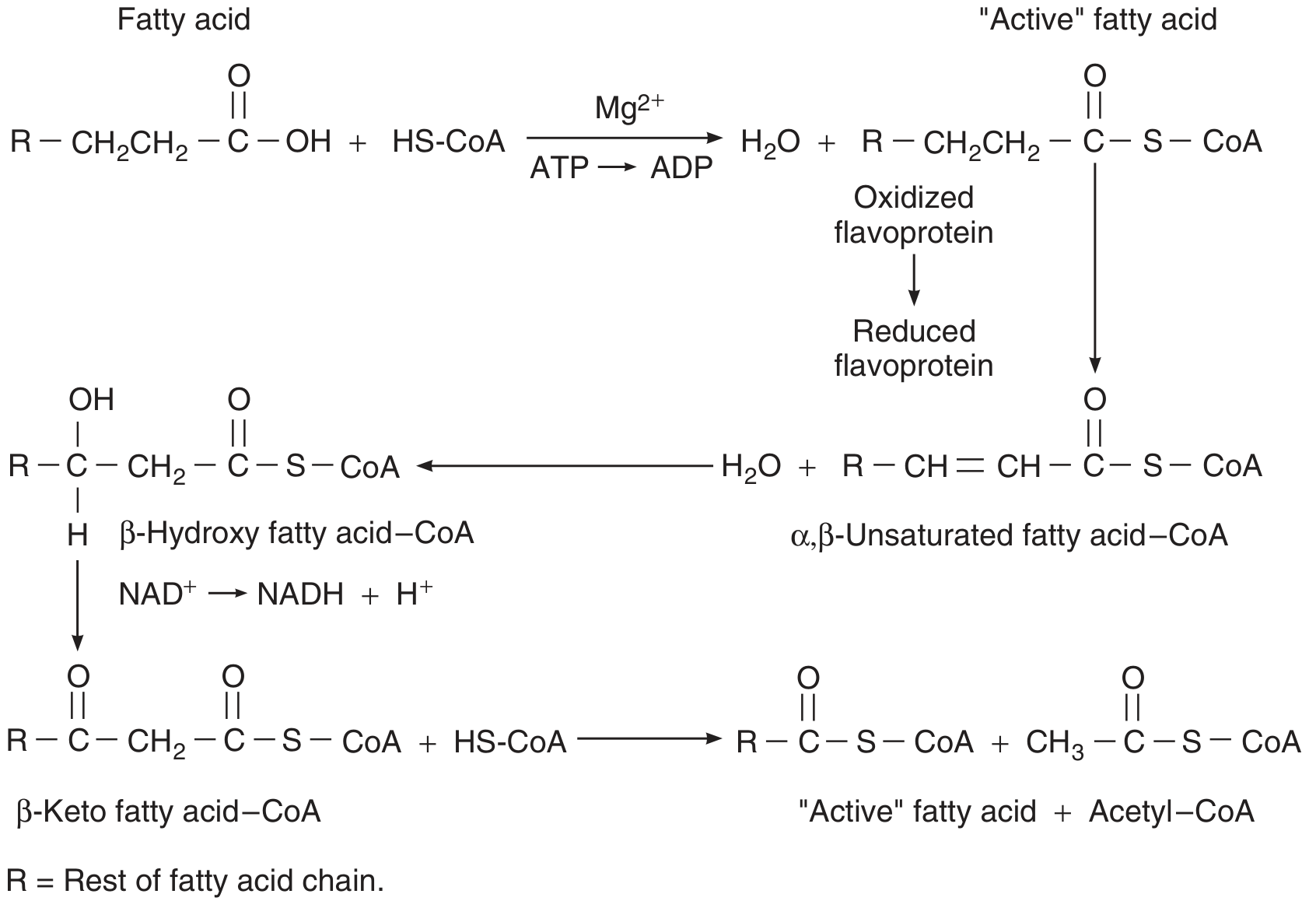
Figure: Fatty acid β-oxidation cycle - Ganong's Review of Medical Physiology
- Oxidation (FAD-dependent): Acyl-CoA → trans-Δ2-Enoyl-CoA (produces FADH₂)
- Hydration: trans-Δ2-Enoyl-CoA → L-3-Hydroxyacyl-CoA
- Oxidation (NAD⁺-dependent): L-3-Hydroxyacyl-CoA → 3-Ketoacyl-CoA (produces NADH)
- Thiolysis: 3-Ketoacyl-CoA + CoA → Acetyl-CoA + Acyl-CoA (shortened by 2C)
Energy Yield Example - Palmitate (16:0)
- 7 cycles of β-oxidation → 8 acetyl-CoA + 7 FADH₂ + 7 NADH
- 8 acetyl-CoA × 10 ATP (via TCA) = 80 ATP
- 7 FADH₂ × 1.5 ATP = 10.5 ATP
- 7 NADH × 2.5 ATP = 17.5 ATP
- Minus 2 ATP for activation = ~106 ATP net
1 mol of a six-carbon FA through the TCA cycle yields 44 mol ATP, compared to 38 mol from 1 mol of glucose (6C). FA are ~2.5× more energy-dense than carbohydrates per gram. - Ganong's Review
Regulation of FA Oxidation
- Malonyl-CoA (the first intermediate in FA synthesis) inhibits CAT I, preventing futile cycling of synthesis and oxidation simultaneously.
- In the fed state: high insulin → high malonyl-CoA → FA synthesis ON, FA oxidation OFF.
- In the fasted state: low insulin → low malonyl-CoA → FA oxidation ON.
5. Ketone Body Synthesis (Ketogenesis)
When acetyl-CoA production exceeds TCA cycle capacity (fasting, starvation, diabetic ketoacidosis), the liver condenses acetyl-CoA units into ketone bodies: acetoacetate, β-hydroxybutyrate, and acetone.
Pathway (in hepatic mitochondria):
- 2 Acetyl-CoA → Acetoacetyl-CoA (thiolase)
- Acetoacetyl-CoA + Acetyl-CoA → HMG-CoA (HMG-CoA synthase)
- HMG-CoA → Acetoacetate + Acetyl-CoA (HMG-CoA lyase)
- Acetoacetate → β-hydroxybutyrate (NADH-dependent) or spontaneously decarboxylates to acetone
Key point: The liver produces ketone bodies but cannot use them (lacks thiophorase/succinyl-CoA:3-ketoacid CoA transferase). The brain, heart, and muscle can oxidize ketone bodies when glucose is scarce, making ketones the brain's major alternative fuel during prolonged starvation.
Normal blood ketone level is ~1 mg/dL. In diabetic ketoacidosis, ketones accumulate because FA oxidation is massively upregulated due to absolute insulin deficiency. - Ganong's Review
6. De Novo Fatty Acid Synthesis
Occurs primarily in the liver and lactating mammary glands (cytosol). This anabolic process is reductive and endergonic, requiring ATP and NADPH.
Step 1 - Acetyl-CoA Transport to Cytosol (Citrate Shuttle)
Mitochondrial acetyl-CoA cannot cross the inner membrane directly. It condenses with OAA → citrate, which is exported to the cytosol. ATP-citrate lyase cleaves citrate back to OAA + acetyl-CoA in the cytosol. This shuttle is activated when ATP levels are high (citrate accumulates when isocitrate dehydrogenase is inhibited by high ATP). - Lippincott Biochemistry, 8e
Step 2 - Carboxylation of Acetyl-CoA to Malonyl-CoA (Rate-Limiting Step)
Acetyl-CoA carboxylase (ACC) - requires biotin (vitamin B7) and ATP:
Acetyl-CoA + HCO₃⁻ + ATP → Malonyl-CoA + ADP + Pi
- ACC is activated by citrate (allosteric) and by insulin-stimulated dephosphorylation.
- ACC is inhibited by palmitoyl-CoA (product feedback), glucagon/epinephrine (via cAMP → AMPK phosphorylation), and malonyl-CoA availability drives chain elongation.
Step 3 - The FAS Complex (Fatty Acid Synthase)
In animals, FAS is a multifunctional enzyme dimer in the cytosol. Using malonyl-CoA as the 2-carbon donor, the growing chain is elongated in a cycle of:
Condensation → Reduction (NADPH) → Dehydration → Reduction (NADPH)
After 7 cycles starting from one acetyl-CoA primer + 7 malonyl-CoA:
- Product: Palmitoyl-CoA (16:0) is released by thioesterase
- Requires: 7 ATP (for malonyl-CoA formation) + 14 NADPH
Elongation beyond 16C and desaturation occur in the ER and mitochondria.
Regulation of Fatty Acid Synthesis
| Signal | Effect on ACC | Effect on FA synthesis |
|---|---|---|
| Insulin (fed state) | Activates (dephosphorylation) | ↑ |
| Glucagon/Epinephrine | Inhibits (AMPK phosphorylation) | ↓ |
| Citrate | Allosteric activation | ↑ |
| Palmitoyl-CoA | Allosteric inhibition | ↓ |
| AMP/AMPK | Phosphorylates ACC (inactive) | ↓ |
7. Cholesterol Metabolism
Structure
Cholesterol is a sterol - four fused hydrocarbon rings (A-D, steroid nucleus), an 8-carbon branched chain at C17, and a hydroxyl group at C3. It is very hydrophobic.
Functions
- Structural component of all cell membranes (modulates fluidity)
- Precursor for bile acids, steroid hormones (glucocorticoids, mineralocorticoids, sex hormones), and vitamin D
De Novo Biosynthesis (Mevalonate Pathway)
All 27 carbons of cholesterol come from acetyl-CoA. Key steps:
- 3 Acetyl-CoA → HMG-CoA (in cytosol)
- HMG-CoA → Mevalonate - catalyzed by HMG-CoA reductase (rate-limiting, ER-bound, target of statins)
- Mevalonate → Farnesyl pyrophosphate (via isoprene intermediates)
- 2 × Farnesyl-PP → Squalene → Lanosterol → Cholesterol
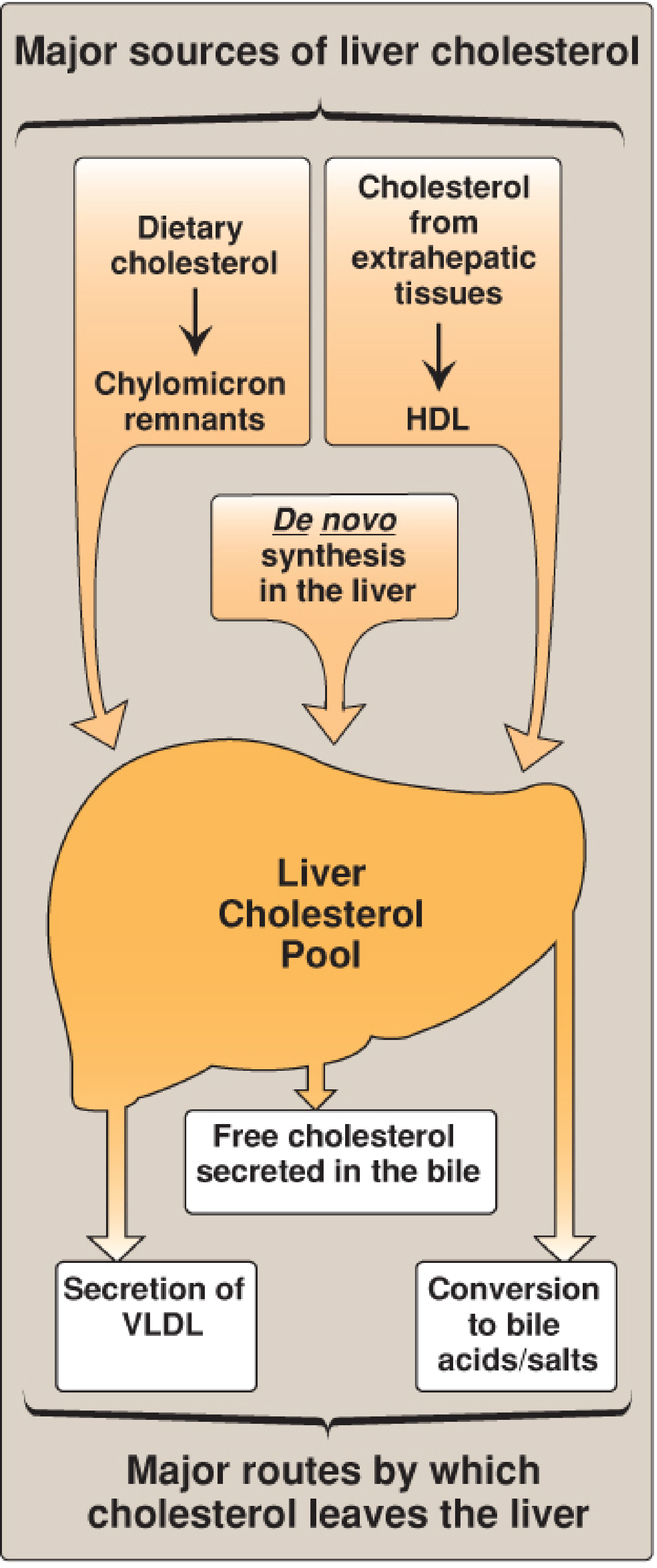
Figure: Major sources of liver cholesterol (influx) and routes by which cholesterol leaves the liver (efflux) - Lippincott Biochemistry, 8e
Regulation of HMG-CoA Reductase
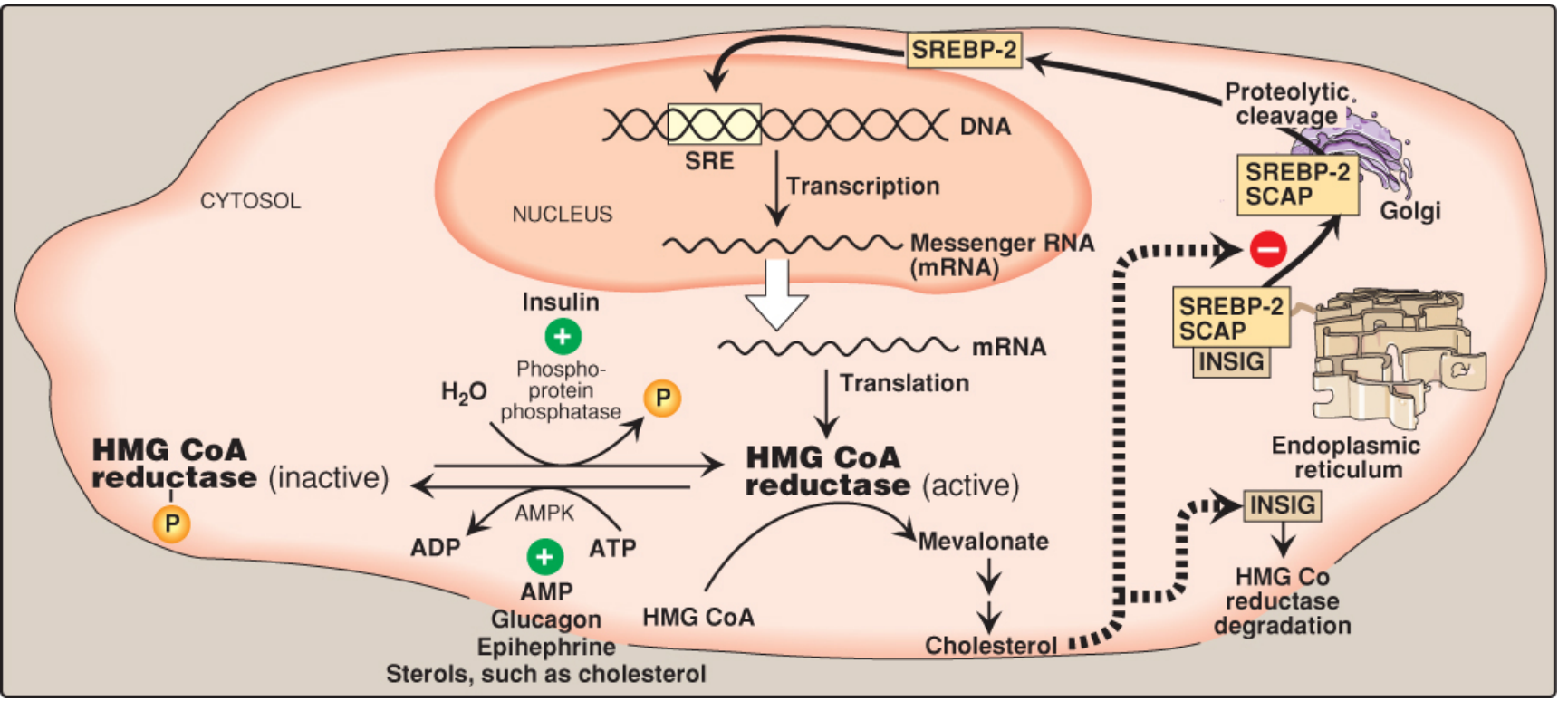
Figure: Regulation of HMG-CoA reductase by sterols (SREBP-2/SCAP/INSIG pathway), phosphorylation (AMPK), and hormones - Lippincott Biochemistry, 8e
Four regulatory mechanisms:
-
Sterol-dependent transcription (SREBP-2 pathway): When ER cholesterol is low, SCAP escorts SREBP-2 to the Golgi, where it is cleaved. Active SREBP-2 enters the nucleus and upregulates HMG-CoA reductase transcription. When sterols are high, SCAP-SREBP is retained in the ER by INSIG proteins.
-
Sterol-accelerated enzyme degradation: High ER sterols promote binding of reductase to INSIG → ubiquitination → proteasomal degradation.
-
Phosphorylation/dephosphorylation: AMPK phosphorylates and inactivates the reductase. Insulin favors the active dephosphorylated form; glucagon and epinephrine favor the inactive phosphorylated form.
-
Hormonal regulation: Insulin activates, glucagon/epinephrine inhibits.
Because AMPK is activated by AMP, cholesterol synthesis is decreased when ATP availability is low - the same condition that inhibits FA synthesis. - Lippincott Biochemistry, 8e
Cholesterol Intestinal Absorption
Niemann-Pick C1-like 1 (NPC1L1) protein mediates intestinal cholesterol uptake - this is the target of ezetimibe. Plant sterols (phytosterols) compete with cholesterol for absorption but are actively effluxed back into the intestinal lumen by ABCG5/8. Defective ABCG5/8 causes sitosterolemia.
Bile Acid Synthesis
Cholesterol is converted to primary bile acids (cholic acid, chenodeoxycholic acid) in the liver. The rate-limiting enzyme is 7α-hydroxylase (CYP7A1), which is:
- Upregulated by cholesterol (via LXR nuclear receptor)
- Downregulated by bile acids (via FXR nuclear receptor)
Bile acids are conjugated with glycine or taurine before secretion, making them more amphipathic and effective as emulsifiers at intestinal pH.
8. Integration of Lipid Metabolism
| State | Dominant Pathway |
|---|---|
| Fed/high insulin | FA synthesis, TAG synthesis, cholesterol synthesis; LPL active in adipose |
| Fasted/low insulin | FA mobilization from adipose (HSL), β-oxidation, ketogenesis in liver |
| Prolonged starvation | Ketone bodies serve as major brain fuel; gluconeogenesis from glycerol |
| Exercise | LPL active in muscle; FA oxidation increases |
| Diabetic ketoacidosis | Uncontrolled FA release → massive β-oxidation → ketosis; metabolic acidosis |
Key regulatory enzymes and their control:
| Enzyme | Activators | Inhibitors |
|---|---|---|
| ACC (FA synthesis) | Citrate, insulin | Palmitoyl-CoA, AMPK, glucagon |
| HMG-CoA reductase (cholesterol synthesis) | Insulin, dephosphorylation | Sterols, AMPK, glucagon |
| CAT I (FA transport into mitochondria) | Low malonyl-CoA | Malonyl-CoA |
| Hormone-sensitive lipase (TAG hydrolysis in adipose) | Glucagon, epinephrine (cAMP) | Insulin |
| Lipoprotein lipase (TAG hydrolysis in blood) | Insulin, ApoC-II | Fasting, ApoC-III |
Sources:
- Lippincott's Illustrated Reviews: Biochemistry, 8th ed. - Chapters 15, 16, 18
- Ganong's Review of Medical Physiology, 26th ed. - Chapter 1 (Fatty Acid Oxidation & Synthesis)
- Guyton and Hall Textbook of Medical Physiology - Chapter 69 (Lipid Metabolism)
This is a shared conversation. Sign in to Orris to start your own chat.