Neural And Chemical Regulation Of Respiration
Neural and Chemical Regulation of Respiration
1. The Respiratory Center
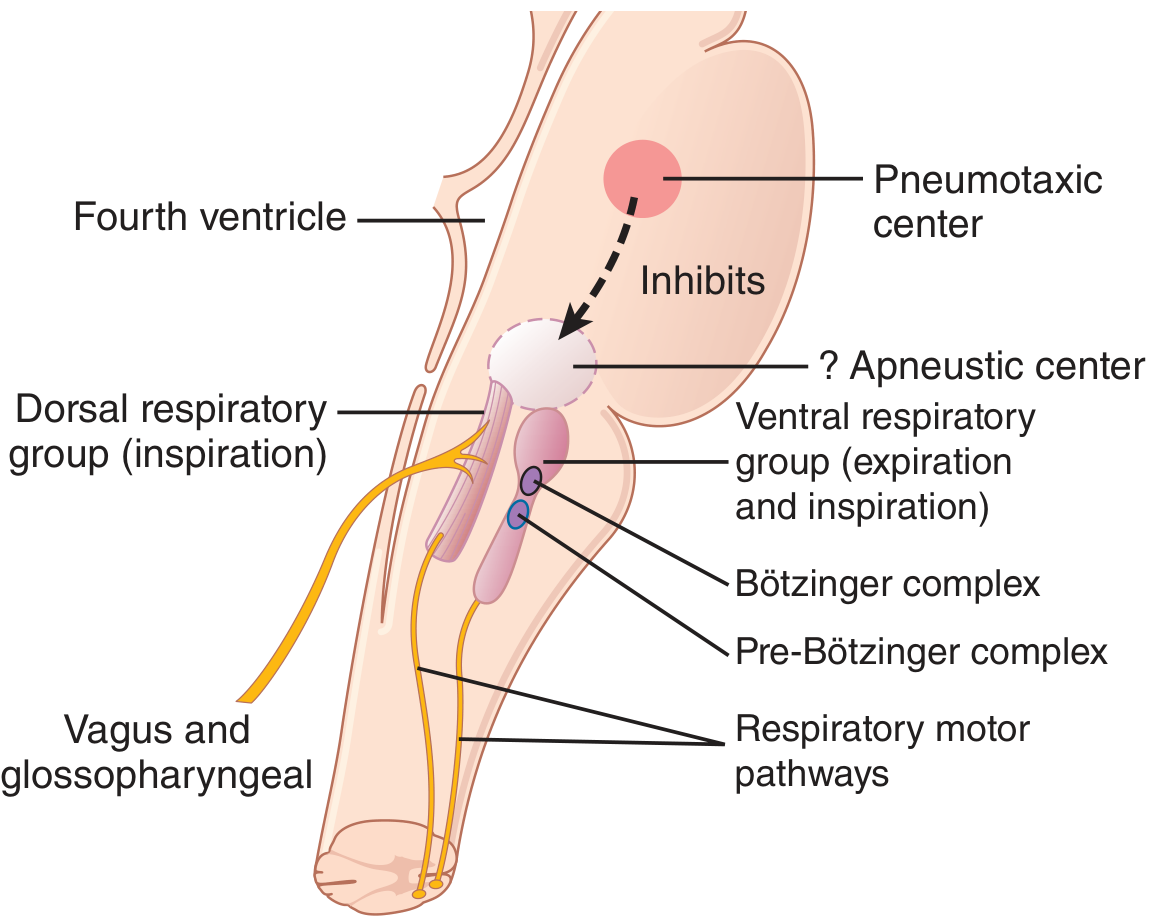
A. Dorsal Respiratory Group (DRG)
- Location: Nucleus of the Tractus Solitarius (NTS) in the dorsal medulla
- Primary function: Controls inspiration and the basic respiratory rhythm
- Receives sensory input via the vagus and glossopharyngeal nerves from: peripheral chemoreceptors, baroreceptors, lung receptors, and GI tract receptors
- Even after all peripheral nerve inputs are cut and the brainstem is transected, this group still emits repetitive bursts of inspiratory action potentials, indicating intrinsic rhythmicity
- Rate of increase (faster in heavy breathing)
- Cut-off point (earlier cut-off = shorter inspiration = faster respiratory rate)
B. Pre-Bötzinger Complex
- Located in the rostral ventral respiratory group
- Contains spontaneously-firing pacemaker neurons with voltage-dependent properties
- Projects to both the DRG and VRG
- Considered a key component of the respiratory central pattern generator (CPG) - its removal eliminates respiratory rhythm generation
- Guyton & Hall, p. 532
C. Ventral Respiratory Group (VRG)
- Location: ~5 mm anterior and lateral to DRG, in the nucleus ambiguus (rostral) and nucleus retroambiguus (caudal)
- Functions differently from DRG:
- Active in both inspiration and expiration (unlike DRG which is purely inspiratory)
- During normal quiet breathing (eupnea), most VRG neurons are inactive - the DRG and pre-Bötzinger complex handle quiet breathing
- When high ventilation is demanded (heavy exercise), the VRG is recruited as an "overdrive mechanism" - it provides powerful expiratory signals to the abdominal muscles
- Guyton & Hall, p. 532
D. Pneumotaxic Center (Pontine Respiratory Group)
- Location: Nucleus parabrachialis, dorsal upper pons
- Primary role: Controls the "switch-off" point of the inspiratory ramp - i.e., it limits the duration of inspiration
- Strong pneumotaxic signal → short inspiration (as little as 0.5 sec) → fast respiratory rate (30-40 breaths/min)
- Weak pneumotaxic signal → prolonged inspiration (up to 5+ sec) → slow respiratory rate (3-5 breaths/min)
- The pneumotaxic center does NOT initiate breathing; it modulates it
- Guyton & Hall, p. 532
E. Apneustic Center (Lower Pons)
- An excitatory center in the lower pons that drives the inspiratory area
- The pneumotaxic center inhibits it
- If the pneumotaxic center is destroyed and the vagi are cut, the apneustic center causes apneusis - prolonged, gasping inspirations
2. Lung Reflexes (Neural Feedback)
Hering-Breuer Inflation Reflex
- Stretch receptors in bronchial and bronchiolar walls detect lung over-inflation
- Signals travel via the vagus nerves to the DRG
- Effect: Switches off the inspiratory ramp ("stops further inspiration") - similar to the pneumotaxic center's action
- In humans, this reflex is activated only when tidal volume exceeds ~1.5 L (>3x normal)
- It is primarily a protective mechanism against excess lung inflation, not a major regulator of normal breathing
- Guyton & Hall, p. 533
3. Chemical Control of Respiration
- Central chemoreceptors (respond to CO₂/H⁺ in CSF)
- Peripheral chemoreceptors (respond primarily to arterial PO₂, and also CO₂/H⁺)
A. Central (Brain) Chemoreceptors - CO₂ and H⁺
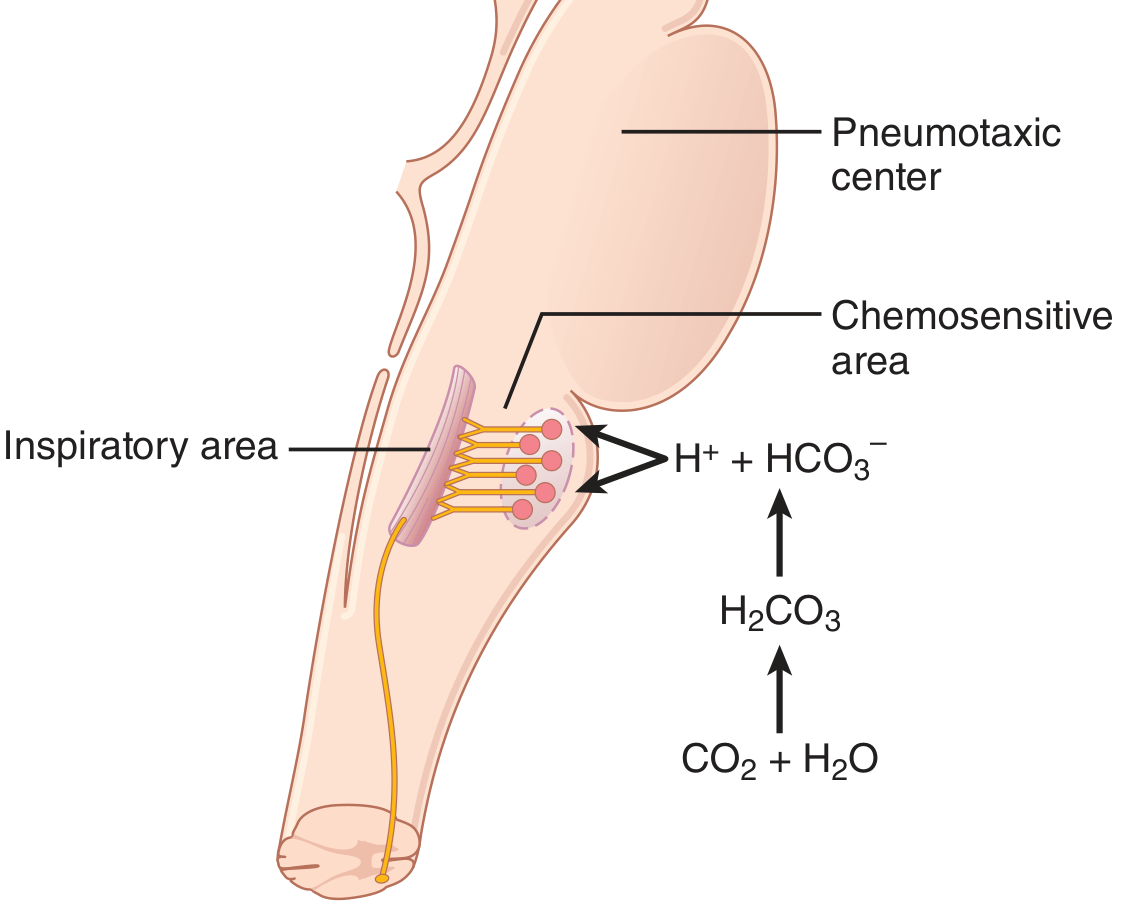
- CO₂ crosses the BBB freely (unlike H⁺)
- In the CSF, CO₂ reacts with water: CO₂ + H₂O → H₂CO₃ → H⁺ + HCO₃⁻
- This locally generated H⁺ directly stimulates the chemosensitive neurons
- Result: Blood CO₂ rise is a far more potent respiratory stimulant than blood H⁺ rise (~7x more potent via central route)
- Guyton & Hall, p. 534
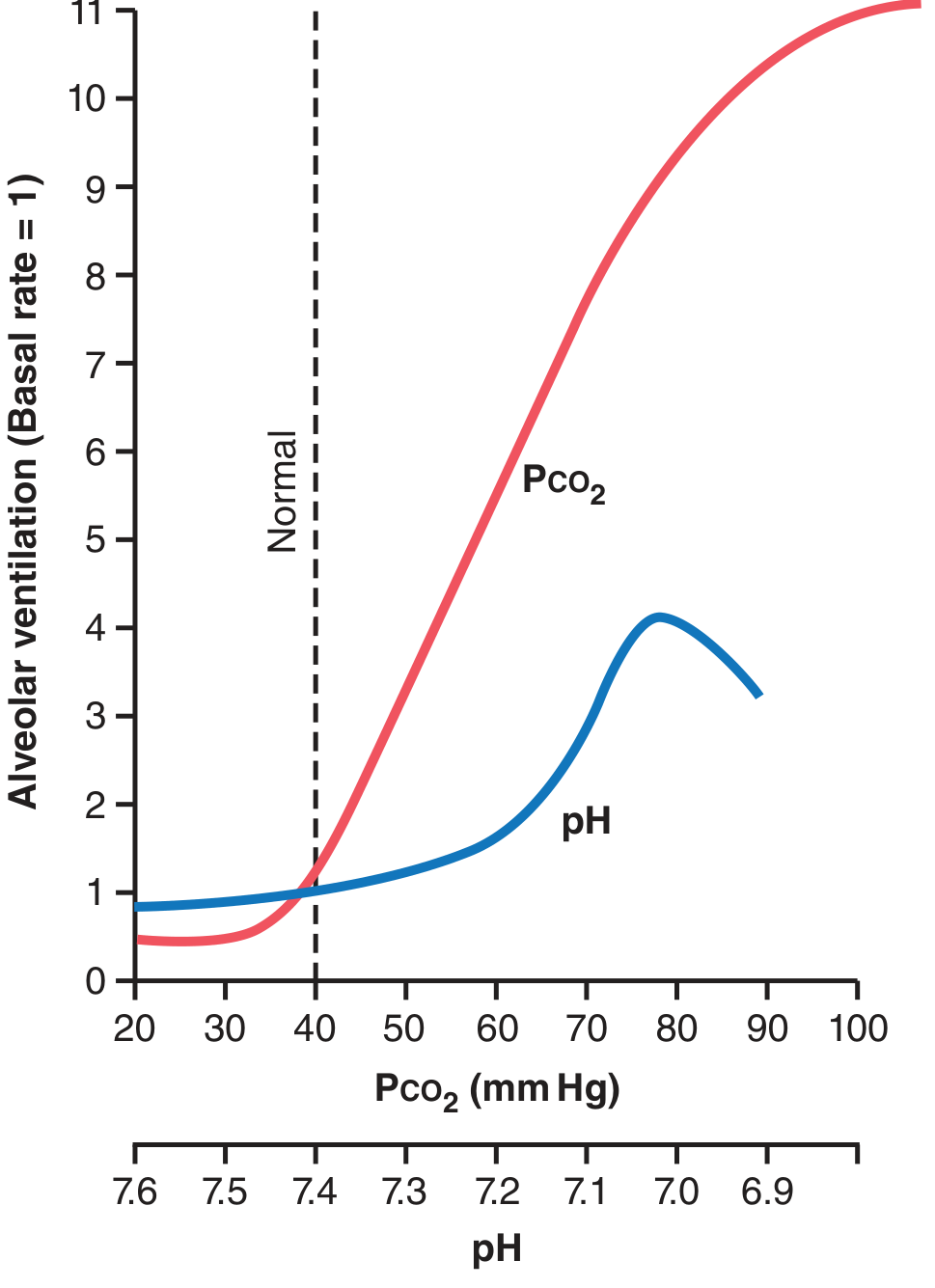
- The CO₂ response is steep in the 35-75 mmHg range
- pH change in the normal range (7.3-7.5) produces <10% of the ventilatory effect of CO₂
B. Peripheral Chemoreceptors - O₂, CO₂, H⁺
- Carotid bodies (bilateral, at the bifurcation of the common carotid arteries) - the dominant peripheral chemoreceptors
- Afferent fibers → Hering's nerve → glossopharyngeal nerve (CN IX) → DRG of medulla
- Aortic bodies (along the arch of the aorta)
- Afferent fibers → vagus nerve (CN X) → DRG of medulla
- A few chemoreceptors also exist along thoracic and abdominal arteries
- Specialized glomus (type I) cells function as the actual O₂ sensors
- When arterial PO₂ falls below ~60 mmHg:
- K⁺ channels close → cell depolarizes
- Ca²⁺ channels open → cytosolic Ca²⁺ rises
- Neurotransmitter release (ATP is likely most important) → activates afferent fibers → respiratory center stimulated
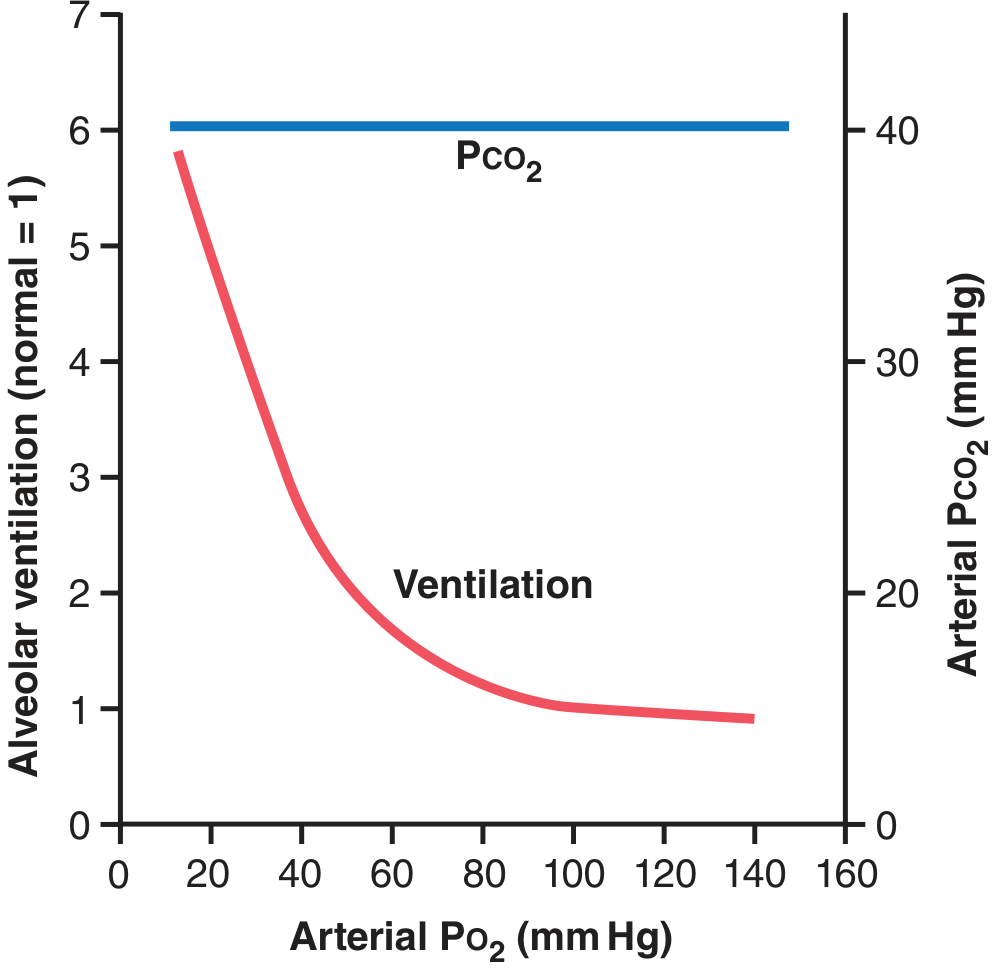
- Above 100 mmHg PO₂: almost no ventilatory effect
- At 60 mmHg PO₂: ventilation approximately doubles
- Very low PO₂: ventilation can increase up to 5-fold
- Conclusion: The peripheral chemoreceptor O₂ response only kicks in significantly below 60-80 mmHg PO₂. At sea level, CO₂/H⁺ responses dominate ventilatory control.
4. Summary: The Three Chemical Regulators
| Stimulus | Primary Receptor | Mechanism | Potency |
|---|---|---|---|
| ↑ PCO₂ | Central (RTN/ventrolateral medulla) | CO₂ → CSF H⁺ → stimulates neurons | Most potent normal regulator |
| ↓ pH / ↑ H⁺ | Central (direct) + peripheral | Direct H⁺ excitation; limited BBB crossing | Less potent than CO₂ alone |
| ↓ PO₂ | Peripheral (carotid/aortic bodies) | Glomus cell K⁺ channel closure → depolarization | Only significant below ~60-80 mmHg |
5. Regulation During Exercise
- The brain transmits collateral (feed-forward) signals from the motor cortex to the brainstem respiratory center simultaneously with signals to the exercising muscles
- This neurogenic drive precedes any detectable chemical change in the blood
- Neural (feedforward) stimuli set ventilation at approximately the correct level
- Chemical feedback then fine-tunes it:
- If too much ventilation → PCO₂ falls → less chemical drive (slight inhibition)
- If too little ventilation → PCO₂ rises / PO₂ falls → chemical drive boosts ventilation
- The result is nearly perfect matching of ventilation to metabolic demands
- Guyton & Hall, p. 537
6. Higher Centers and Other Inputs
- Cerebral cortex: Voluntary override of breathing (breath-holding, deliberate hyperventilation, speech, playing wind instruments)
- Hypothalamus: Modulates respiration in response to fever, emotional states
- Limbic system: Emotional breathing patterns (crying, laughing, sighing)
- J-receptors (juxtacapillary receptors): Located in the alveolar walls near capillaries; stimulated by pulmonary edema or congestion → rapid, shallow breathing (tachypnea)
- Irritant receptors: In airway epithelium; respond to noxious stimuli → cough, bronchoconstriction
7. Acclimatization to Altitude (Chronic Hypoxia)
- The initial peripheral chemoreceptor-driven hyperventilation blows off CO₂
- Normally, the resulting low PCO₂ and high pH would inhibit further increase in ventilation
- But over 2-3 days, the respiratory center loses ~80% of its sensitivity to CO₂/H⁺ (the kidneys excrete HCO₃⁻, normalizing CSF pH)
- This allows low O₂ to drive ventilation to much higher levels
- Alveolar ventilation can increase 400-500% after 2-3 days (vs. only ~70% acutely)
- Guyton & Hall, p. 536
- Guyton and Hall Textbook of Medical Physiology, Chapter 42, pp. 531-538
- Costanzo Physiology, 7th Edition, Chapter 5 (Respiratory Physiology)
- Ganong's Review of Medical Physiology, 26th Edition, Chapter on Pulmonary Physiology
General flowchart photo
A clean, detailed medical flowchart showing the Neural and Chemical Regulation of Respiration. Include: Respiratory Center (medulla + pons) at the top with three branches: Dorsal Respiratory Group (inspiration), Ventral Respiratory Group (inspiration + expiration), and Pneumotaxic Center (limits inspiration). Show the Pre-Bötzinger Complex as the rhythm generator. Below, show two pathways: (1) Central Chemoreceptors detecting CO2/H+ in CSF with the equation CO2 + H2O → H2CO3 → H+ + HCO3-, and (2) Peripheral Chemoreceptors (carotid bodies via CN IX, aortic bodies via CN X) detecting low PO2. Show arrows from both chemoreceptors feeding back to the respiratory center. Include the Hering-Breuer reflex arc from lung stretch receptors via vagus nerve. At the bottom, show the effectors: Diaphragm and intercostal muscles producing Inspiration and Expiration. Use color coding: blue for neural pathways, red for chemical pathways, green for reflex arcs. Professional medical illustration style with clear labels and arrows.
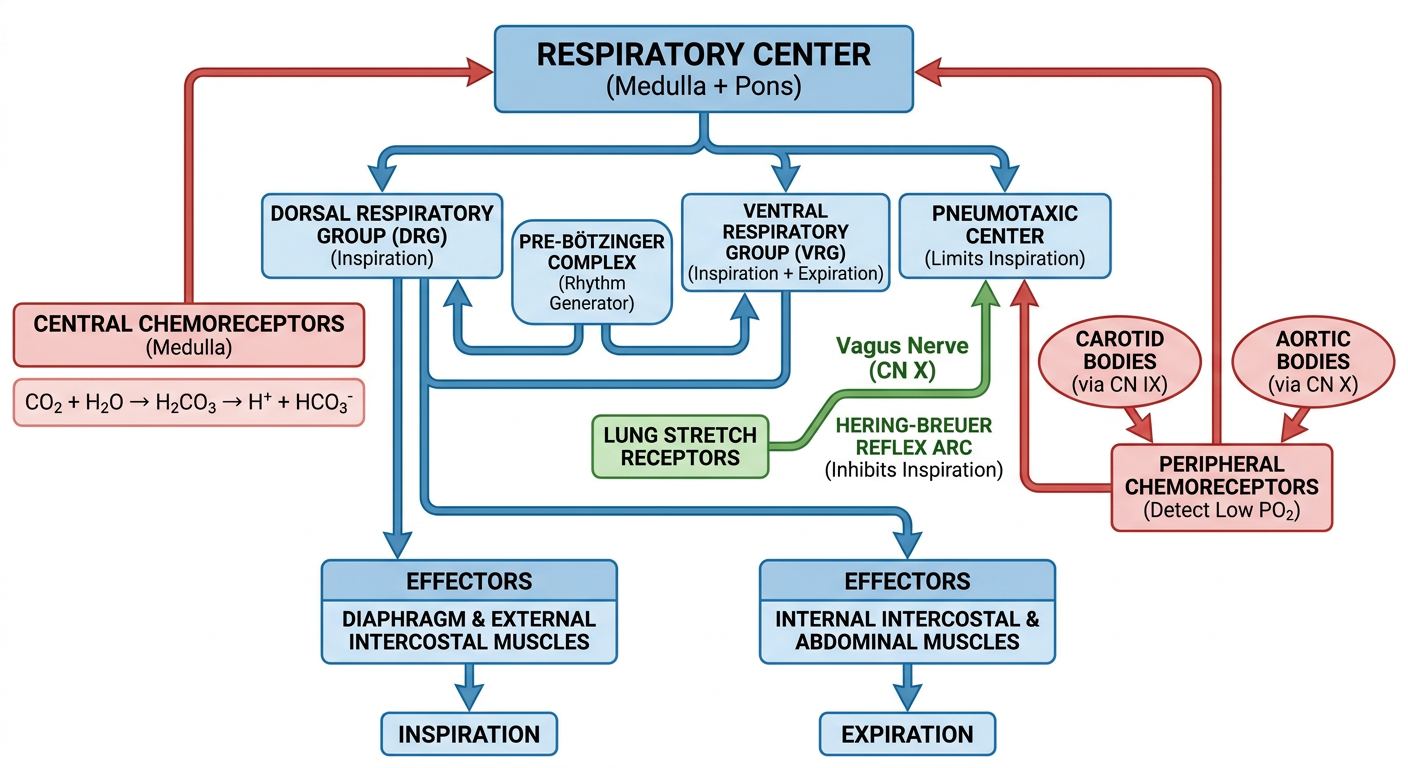
- Blue pathways - Neural control (respiratory center groups, Pre-Bötzinger rhythm generator)
- Red pathways - Chemical control (central CO₂/H⁺ detection, peripheral O₂ detection)
- Green pathways - Reflex arcs (Hering-Breuer inflation reflex via vagus)