Theories of colour vision
Reading File
Finding Sources
Searching the Web
theories of colour vision trichromatic opponent process dual process physiology
Reading File
Reading File
Reading File
Reading File
Loading Image
Finding Sources
I have all the key information I need. Let me now compose the comprehensive answer.
Theories of Colour Vision
Colour vision has been explained by three major theories, each operating at a different level of the visual system. Modern understanding combines all three into a unified, two-stage model.
1. Trichromatic Theory (Young-Helmholtz Theory)
Proposed by: Thomas Young (1802) and later refined by Hermann von Helmholtz (1850s).
Core idea: The retina contains three types of cone photoreceptors, each maximally sensitive to one of three primary colours - red, green, and blue. Any perceived colour results from the combined ratio of stimulation across these three cone types.
The Three Cone Types (S, M, L)
| Cone Type | Common Name | Peak Absorption | Wavelength Range |
|---|---|---|---|
| S (Short-wave) | Blue-sensitive | ~430 nm (violet-blue) | 400-500 nm |
| M (Medium-wave) | Green-sensitive | ~530 nm (green) | 450-630 nm |
| L (Long-wave) | Red-sensitive | ~560 nm (yellow-red) | 500-700 nm |
The diagram below from Guyton shows the spectral absorption curves and the ratios of cone stimulation by different colours:
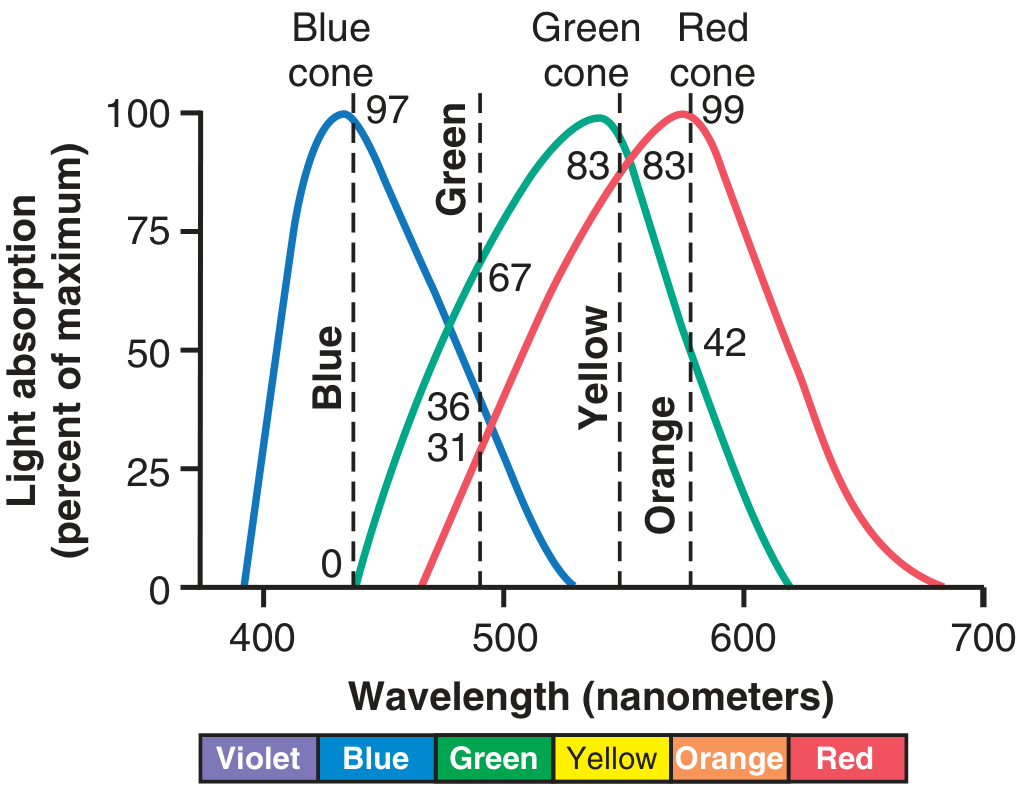
Colour Interpretation by Ratio
The nervous system interprets colour by the ratio of stimulation across cone types, not by absolute values. Examples (from Guyton & Hall):
- Orange (580 nm): Red cones 99, Green 42, Blue 0 → ratio 99:42:0
- Blue (450 nm): Red 0, Green 0, Blue 97 → ratio 0:0:97
- Yellow: Red 83, Green 83, Blue 0 → ratio 83:83:0
- Green: ratio 31:67:36
- White: approximately equal stimulation of all three
Genetic Basis
- Rhodopsin gene: chromosome 3
- Blue (S) cone pigment gene: chromosome 7
- Green (M) and Red (L) cone pigment genes: tandemly arranged on the q arm of the X chromosome - explaining why red-green colour blindness is X-linked
- M and L opsins share 96% amino acid homology with each other, but only ~43% with the S-cone opsin and ~41% with rhodopsin
(Ganong's Review of Medical Physiology, p. 209)
2. Opponent Process Theory (Hering's Theory)
Proposed by: Ewald Hering (1878).
Core idea: Colour perception beyond the cones is processed by antagonistic (opponent) pairs. Specific ganglion cells and LGN neurons respond by being excited by one colour and inhibited by its opponent.
The Three Opponent Channels
| Channel | Opponent Pair | Mechanism |
|---|---|---|
| Red-Green | Red vs. Green | L-cone signals minus M-cone signals |
| Blue-Yellow | Blue vs. Yellow | S-cone signals minus (L+M) cone sum |
| Black-White (Luminance) | Light vs. Dark | Sum of L+M cone responses |
This explains several phenomena that trichromatic theory alone cannot:
- Afterimages: Staring at red produces a green afterimage (opponent rebound)
- Simultaneous contrast: A red object appears pale/white when surrounded by red light
- Why reddish-green and bluish-yellow do not exist as perceptual colours - opponents cancel each other
As Ganong states: "Color is mediated by ganglion cells that subtract or add input from one type of cone to input from another type." - Ganong's Review of Medical Physiology, p. 210
3. Dual Process (Stage) Theory - The Modern Synthesis
Proposed by: Leo Hurvich and Dorothea Jameson (1957), building on earlier work.
This reconciles both previous theories by showing they operate at different stages of the visual pathway:
Stage 1 - Retinal Level (Trichromatic)
The three cone types (S, M, L) independently absorb light. This is the Young-Helmholtz stage.
Stage 2 - Retinal/LGN Level (Opponent Process)
Ganglion cells and lateral geniculate nucleus (LGN) neurons combine and compare cone signals into three opponent channels:
- Red-Green pathway - signals the difference between L and M cone responses
- Blue-Yellow pathway - signals the difference between S cone response and the sum of L+M responses
- Luminance pathway - signals the sum of L and M cone responses
These project to the blobs and the deep portion of layer 4C of V1 (primary visual cortex). From V1 blobs and layer 4, colour information is projected to area V8 of the visual cortex (the dedicated colour processing area). Lesions of V8 produce achromatopsia (loss of colour vision with preserved form perception).
(Ganong's Review of Medical Physiology, p. 210)
Clinical Relevance: Colour Blindness
Defects in trichromatic theory explains colour blindness:
| Term | Meaning |
|---|---|
| Trichromat | Normal - all three cone systems present |
| Dichromat | Only two cone systems; cannot distinguish some colours |
| Monochromat | Only one cone system; matches spectrum by intensity only |
| Protanopia | Missing red (L) cones |
| Deuteranopia | Missing green (M) cones |
| Tritanopia | Missing blue (S) cones (rare, no sex predilection) |
| Achromatopsia | Cortical colour blindness (V8 lesion) |
Prevalence: Abnormal colour vision affects ~8% of white males and ~0.4% of white females (mostly red-green, X-linked recessive). Tritanopia is rare with no sex selectivity.
A notable pharmacological cause: Sildenafil (Viagra) inhibits the retinal form of phosphodiesterase, causing transient blue-green colour weakness as a side effect.
(Ganong's Review of Medical Physiology, 26th Ed.; Guyton & Hall Textbook of Medical Physiology)
Summary Table
| Theory | Proposed By | Site | Mechanism |
|---|---|---|---|
| Trichromatic | Young-Helmholtz | Retina (cones) | 3 cone types with overlapping spectral sensitivities; colour = ratio of stimulation |
| Opponent Process | Hering | Ganglion cells / LGN | Opponent pairs (R-G, B-Y, Black-White); subtraction of cone signals |
| Dual Process | Hurvich & Jameson | Both stages combined | Trichromatic at cones → Opponent processing at post-receptoral level → Cortical integration in V8 |
The dual-process model is the currently accepted framework - trichromatic encoding at the receptor level feeds into opponent-channel processing at the neural level, ultimately producing the rich spectrum of colour experience in the visual cortex.
This is a shared conversation. Sign in to Orris to start your own chat.