Q2.What are the different type of cell junction? Explain with help of diagram. Q3. Define resting membrane potential and genesis of RMP in detail, with special emphasis on Gibbs Donnan membrane equilibrium. [1] [1] [1] [1] (1] [10]
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Reading File
Loading Image
Loading Image
Loading Image
Loading Image
Reading File
I now have all the information needed for comprehensive answers to both questions. Let me compose the full response.
Q2. Types of Cell Junctions
Cell junctions are specialized membrane-associated structures that mediate adhesion, communication, and occlusion between adjacent cells. They are especially prominent in epithelial tissues. There are four major types, often arranged in a characteristic apico-basal order called the junctional complex.
Diagram - Junctional Complex of Epithelial Cells
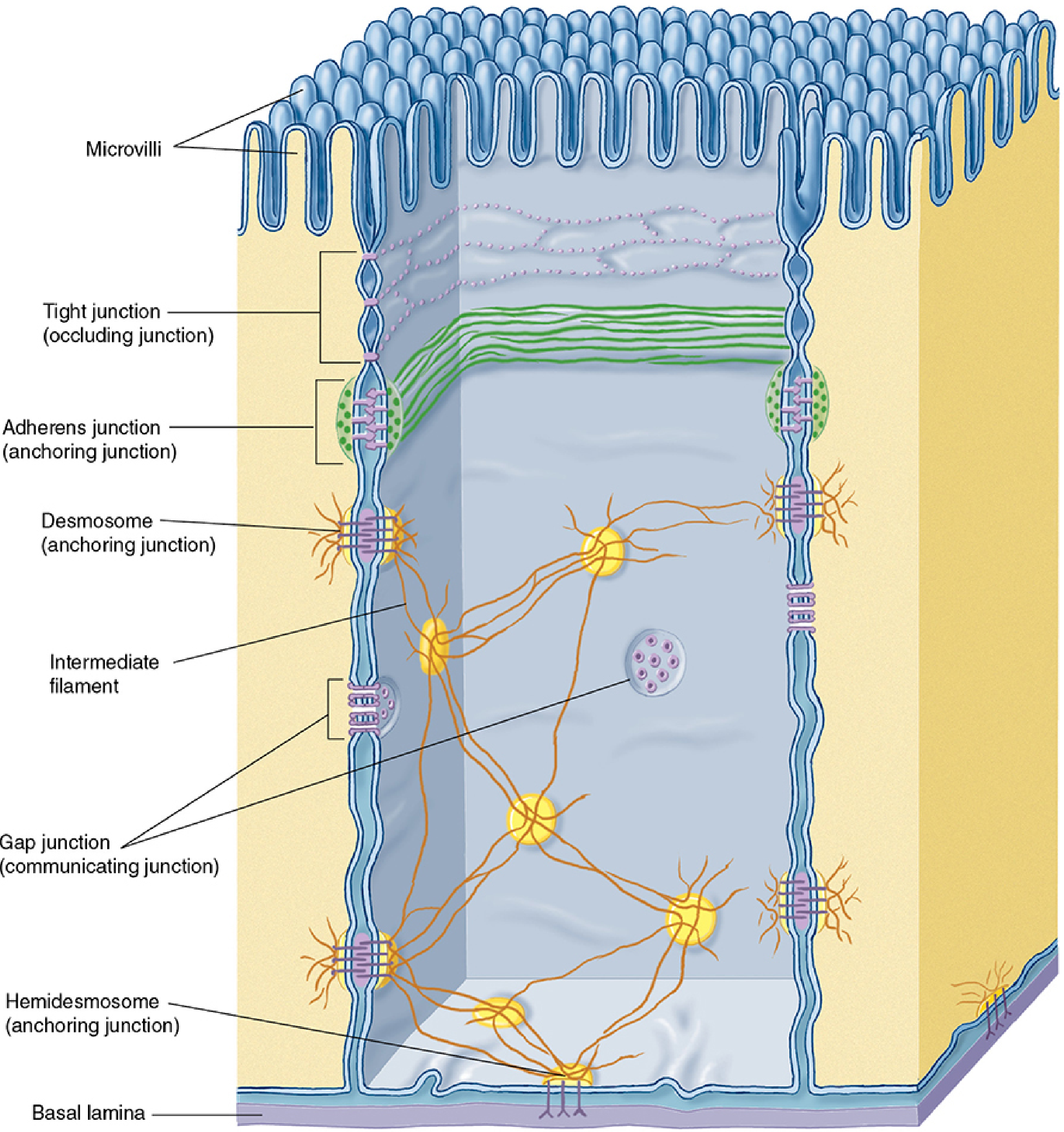
Figure: Junctional complexes of epithelial cells, from Junqueira's Basic Histology
1. Tight Junction (Zonula Occludens) - Occluding Junction
Location: Most apical junction; forms a continuous belt completely encircling each cell.
Structure:
- Adjacent cell membranes appear fused or very tightly apposed in TEM.
- Formed by transmembrane proteins: claudin and occludin, which interact across the intercellular space.
- On cryofracture: appear as branching strands in the membrane.
Functions:
- Seals the intercellular space, preventing paracellular flow of molecules between cells.
- Forces molecules crossing the epithelium to use the transcellular pathway.
- Acts as a fence - prevents mixing of apical and basolateral membrane lipids/proteins, maintaining cell polarity.
- Epithelia with more fused strands (e.g., urinary bladder lining) are less permeable than those with fewer strands (e.g., proximal renal tubule).
Clinical relevance: Clostridium perfringens enterotoxin binds claudin and disrupts tight junctions, causing food poisoning. Helicobacter pylori targets ZO-1 (a tight junction protein) in the stomach.
2. Adherens Junction (Zonula Adherens) - Anchoring Junction
Location: Immediately below the tight junction; also forms a complete belt/band around the cell (zonula = belt).
Structure:
- Transmembrane proteins: E-cadherins (calcium-dependent adhesion molecules).
- Cytoplasmic side linked to actin microfilaments via catenins (alpha, beta, and plakoglobin).
Functions:
- Firmly anchors adjacent cells to each other.
- Stabilizes and strengthens the occluding tight junction above it.
- Helps maintain tissue integrity under mechanical stress.
3. Desmosome (Macula Adherens) - Anchoring Junction
Location: Spot-like (macula = spot), scattered on lateral surfaces below the adherens junction; does not form a belt.
Structure:
- Disc-shaped plaques on each cell that interlock with the adjacent cell.
- Transmembrane proteins: desmogleins and desmocollins (larger cadherin family members).
- Cytoplasmic side: desmogleins bind plakoglobins → desmoplakins → intermediate filaments (cytokeratin/tonofilaments).
Functions:
- Provides very strong spot-weld adhesion - resists shearing and mechanical forces.
- Acts as a major scaffold for tissue integrity, especially in skin and heart.
Clinical relevance: Pemphigus vulgaris - autoimmune antibodies against desmogleins cause loss of cell-cell adhesion and blistering of the skin and oral mucosa.
Hemidesmosome: A variant that anchors epithelial cells to the underlying basal lamina (extracellular matrix), not to another cell. Contains integrins instead of cadherins.
4. Gap Junction (Nexus) - Communicating Junction
Location: Scattered as patch-like structures on lateral cell surfaces.
Structure:
- Each gap junction is a patch of many connexons in adjacent membranes.
- Each connexon is made of 6 protein subunits called connexins arranged around a central pore (~1.5 nm diameter).
- A pair of abutting connexons from two cells forms a complete channel.
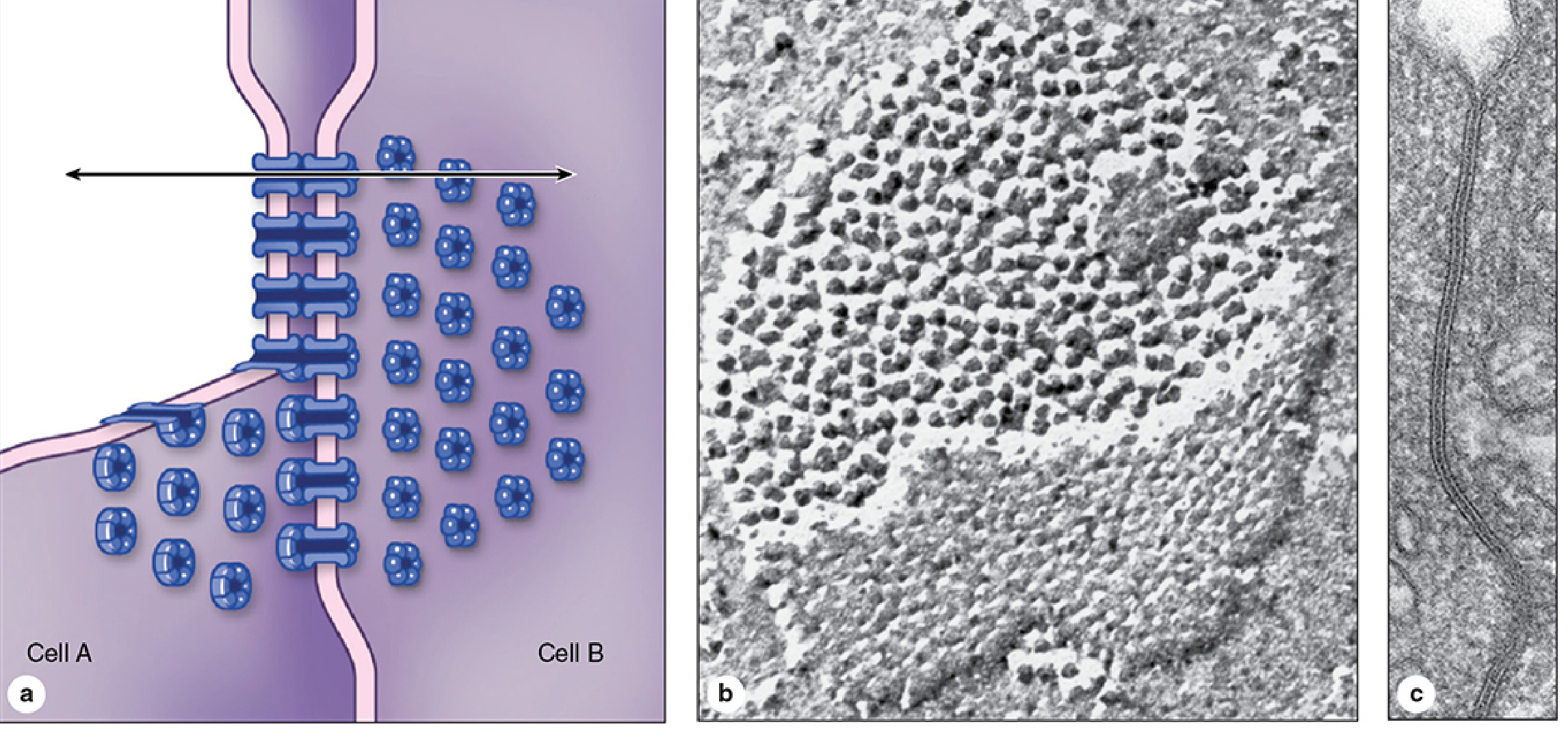
Figure: Gap junction structure - connexons formed by hexameric connexins span both membranes; (b) cryofracture EM showing particle clusters; (c) cross-section view. From Junqueira's Basic Histology
Functions:
- Allows direct intercellular communication - passage of small molecules (ions, cAMP, metabolites, up to ~1 kDa) between adjacent cells.
- No mechanical strength.
- Critical for cardiac muscle synchronization, smooth muscle coordination, and embryonic development.
Summary Table
| Feature | Tight Junction | Adherens Junction | Desmosome | Gap Junction |
|---|---|---|---|---|
| Other name | Zonula occludens | Zonula adherens | Macula adherens | Nexus |
| Shape | Belt (zonula) | Belt (zonula) | Spot (macula) | Patch |
| Key proteins | Claudin, occludin | E-cadherin, catenins | Desmoglein, desmocollin | Connexin (connexons) |
| Cytoskeletal link | - | Actin filaments | Intermediate filaments | - |
| Primary function | Sealing/barrier | Cell-cell adhesion | Strong spot adhesion | Intercellular communication |
| Location | Most apical | Below tight junction | Lateral surface | Lateral surface |
(Source: Junqueira's Basic Histology, 17e)
Q3. Resting Membrane Potential (RMP) and its Genesis
Definition
The resting membrane potential (RMP) is the electrical potential difference that exists across the plasma membrane of an excitable cell (nerve, muscle) when it is not generating an action potential (i.e., at rest). By convention, the intracellular potential is expressed relative to the extracellular potential (which is set at 0 mV).
- In large myelinated nerve fibers: RMP ≈ -70 mV
- In skeletal muscle: RMP ≈ -80 to -90 mV
- In cardiac muscle: RMP ≈ -85 to -90 mV
Prerequisites: Ion Concentration Gradients
The Na⁺-K⁺ ATPase pump establishes and maintains ion concentration gradients across the membrane:
| Ion | Outside (mEq/L) | Inside (mEq/L) |
|---|---|---|
| Na⁺ | 142 | 14 |
| K⁺ | 4 | 140 |
| Cl⁻ | 103 | ~4 |
| Large anions (proteins) | ~0 | ~65 |
The pump transports 3 Na⁺ out for every 2 K⁺ in per cycle, consuming ATP.
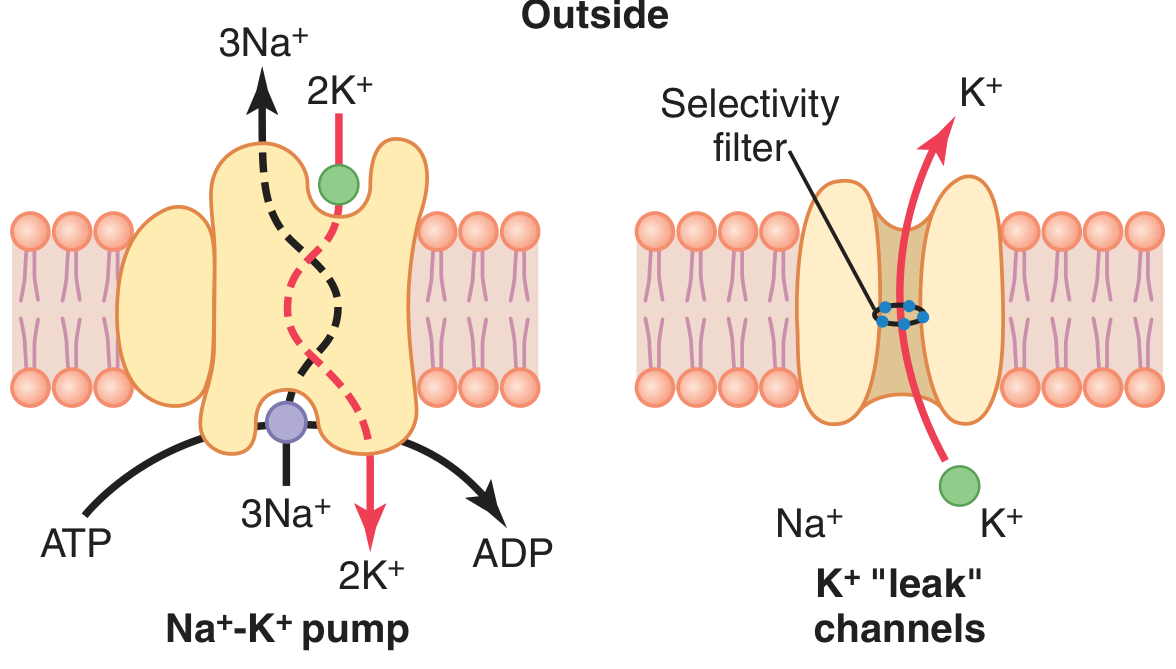
Figure 5.4 from Guyton & Hall: Na⁺-K⁺ pump (left) and K⁺ "leak" channels (right)
Genesis of Resting Membrane Potential
The RMP arises from three interacting factors:
Step 1: Potassium Diffusion Potential
K⁺ is at very high concentration inside the cell (140 mEq/L inside vs. 4 mEq/L outside). K⁺ "leak" channels (tandem pore domain channels) are open at rest, allowing K⁺ to diffuse outward down its concentration gradient. As K⁺ leaves, it carries positive charge out, making the inside progressively more negative. This negative interior then opposes further K⁺ efflux (electrical force). Equilibrium is reached at the Nernst potential for K⁺ = -94 mV.
Nernst equation: E_K = -61 × log([K⁺]i / [K⁺]o) = -61 × log(35) = -94 mV
Step 2: Sodium Diffusion Component
Na⁺ is at higher concentration outside (142 mEq/L) than inside (14 mEq/L). The membrane has small but finite Na⁺ permeability through the K⁺-Na⁺ leak channels (K⁺ permeability is ~100x greater than Na⁺ permeability at rest). Na⁺ tries to diffuse in, with a Nernst potential of +61 mV. Using the Goldman-Hodgkin-Katz (GHK) equation (which weights by relative permeability), the combined effect of K⁺ and Na⁺ diffusion gives approximately -86 mV.
Goldman equation: Em = -61 × log [(PK[K]i + PNa[Na]i) / (PK[K]o + PNa[Na]o)]
Step 3: Na⁺-K⁺ Pump Contribution (Electrogenic)
The pump moves 3 Na⁺ out for every 2 K⁺ in, creating a net outward movement of positive charge. This direct electrogenic effect adds a further -4 mV to the membrane potential. The final resting potential = approximately -90 mV (nerve fibers at full contribution).
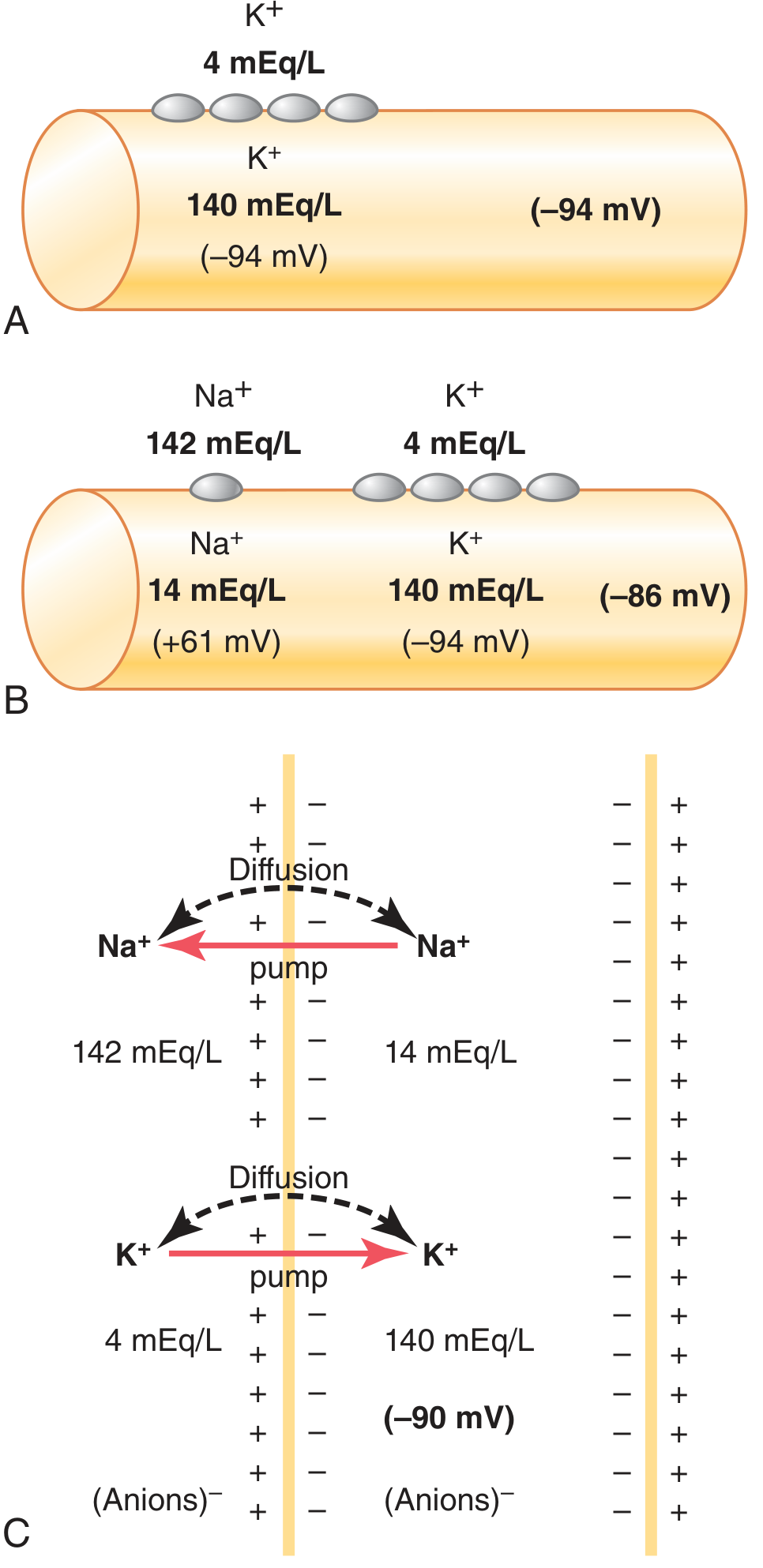
Figure 5.5 from Guyton & Hall: (A) K⁺ diffusion alone = -94 mV; (B) K⁺ + Na⁺ diffusion = -86 mV; (C) + electrogenic Na⁺-K⁺ pump = -90 mV
Gibbs-Donnan Membrane Equilibrium
Concept
The Gibbs-Donnan equilibrium (Donnan equilibrium) describes the unequal distribution of diffusible ions across a membrane that separates a compartment containing non-diffusible (impermeant) charged macromolecules (like intracellular proteins) from one that does not.
Key Principles
A semipermeable membrane separates:
- Outside compartment: Na⁺ = 100 mM, Cl⁻ = 100 mM (no impermeant ions)
- Inside compartment: Na⁺ = 200 mM, Cl⁻ = 50 mM, Y⁻¹⁵⁰ = 1 mM (carries 150 negative charges)
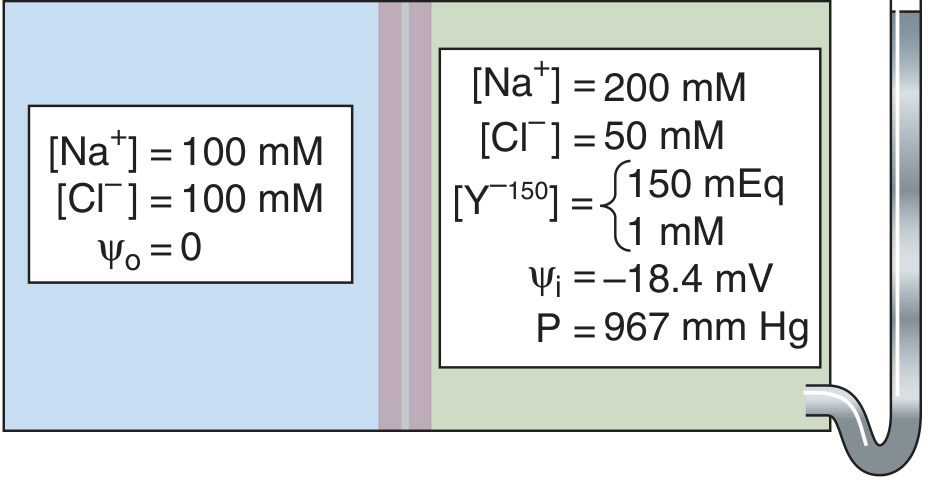
Figure 5-15 from Medical Physiology (Boron & Boulpaep): Gibbs-Donnan equilibrium. Outside: Na⁺=100, Cl⁻=100, ψ=0. Inside: Na⁺=200, Cl⁻=50, Y⁻¹⁵⁰=1 mM, ψᵢ = -18.4 mV, P = 967 mmHg
Mechanism
- Y⁻¹⁵⁰ (impermeant large anion) is trapped inside the cell.
- Cl⁻ moves inward (down its concentration gradient) → generates inside-negative voltage.
- This inside-negative voltage attracts Na⁺ inward.
- At Gibbs-Donnan equilibrium, the membrane potential satisfies the Nernst equation for ALL permeant ions simultaneously:
$$\frac{[Na^+]_o}{[Na^+]_i} = \frac{[Cl^-]_i}{[Cl^-]_o} = r \quad \text{(Donnan ratio)}$$
- The Donnan ratio $r$ is the same for all diffusible monovalent ions. This means:
- Diffusible cations (Na⁺) are more concentrated on the side with the impermeant anion (inside).
- Diffusible anions (Cl⁻) are less concentrated on that same side.
Critical Consequence: Osmotic Imbalance and Cell Swelling
Although ions reach electrochemical equilibrium, there are more total osmotically active particles inside than outside:
- Inside: [Na⁺] + [Cl⁻] = ~250 mOsm
- Outside: [Na⁺] + [Cl⁻] = ~200 mOsm
This 50 mOsm gradient drives water into the cell. In a rigid compartment, this generates a hydrostatic pressure of ~967 mmHg (1.3 atm) to oppose water entry. But cell membranes are not rigid - so in real cells, the Donnan forces would cause progressive water influx, cell swelling, and eventually bursting.
Role of Na⁺-K⁺ Pump in Preventing Donnan-Induced Swelling
The Na⁺-K⁺ ATPase acts as a "pump-leak" steady state to counteract Donnan forces:
- The pump continuously extrudes Na⁺, functionally making the cell impermeable to NaCl.
- NaCl thus acts as a functionally impermeant solute in the extracellular space, offsetting the osmotic effect of intracellular impermeant anions.
- This is not equilibrium but a steady state maintained by active transport.
- If the pump is inhibited, passive entry of Na⁺ exceeds K⁺ efflux → net gain of intracellular cations → depolarization → Cl⁻ entry (follows depolarization) → osmotic swelling → cell lysis.
Relationship to RMP
The Gibbs-Donnan equilibrium contributes to RMP in two ways:
- The impermeant intracellular proteins carry negative charges, attracting K⁺ inward and repelling Cl⁻ outward, tending to make the inside negative.
- It establishes the ionic concentration gradients that the Nernst equation then translates into an equilibrium membrane potential for each ion.
The actual RMP is not the pure Donnan equilibrium potential (which would cause cell swelling), but a steady state maintained by the Na⁺-K⁺ pump that approximates -70 to -90 mV.
Summary of RMP Genesis
Impermeant intracellular anions (proteins, phosphates)
↓
Gibbs-Donnan effect → K⁺ concentrated inside, Na⁺ outside
↓
Na⁺-K⁺ pump maintains concentration gradients (3Na⁺ out / 2K⁺ in)
↓
K⁺ diffuses outward through leak channels → inside becomes -
↓
Small Na⁺ inward leak (Goldman equation) offsets some negativity
↓
Electrogenic pump adds further -4 mV
↓
Net RMP ≈ -70 to -90 mV (inside negative)
(Sources: Guyton & Hall Textbook of Medical Physiology; Costanzo Physiology 7e; Boron & Boulpaep Medical Physiology)
This is a shared conversation. Sign in to Orris to start your own chat.