I have my university exam of 1 st year of subject biochemistry These are the important long answer give me answer of this question "Glycolysis in RBC (Rapoport-Luebering cycle)" and remeber it's a university examination
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Finding Sources
Reading File
Loading Image
Loading Image
I now have comprehensive content from authoritative textbooks with diagrams. Let me write the full university-level answer.
Glycolysis in RBC and the Rapoport-Luebering Cycle
Introduction
Red blood cells (RBCs/erythrocytes) are unique among body cells because they lack a nucleus, mitochondria, and all other membrane-bound organelles. As a result, they cannot carry out oxidative phosphorylation or the TCA cycle. Glucose is their sole source of energy, and glycolysis (Embden-Meyerhof Pathway - EMP) is the only pathway for ATP generation. In addition to standard glycolysis, the RBC possesses a specialized bypass called the Rapoport-Luebering (RL) shunt (cycle), which is unique to erythrocytes and is responsible for generating 2,3-bisphosphoglycerate (2,3-BPG), a key regulator of oxygen delivery to tissues.
Glycolysis in RBC - Overview
Mature RBCs metabolize approximately 90% of their glucose through glycolysis. The overall pathway is the same as in other cells (Embden-Meyerhof Pathway), but it has two important branches unique to erythrocytes:
- Hexose Monophosphate (HMP) Shunt - ~5-10% of glucose, protects against oxidative damage via NADPH
- Rapoport-Luebering Shunt - generates 2,3-BPG to regulate hemoglobin-oxygen affinity
FIGURE 42.1 - Overview of Erythrocyte Metabolism:
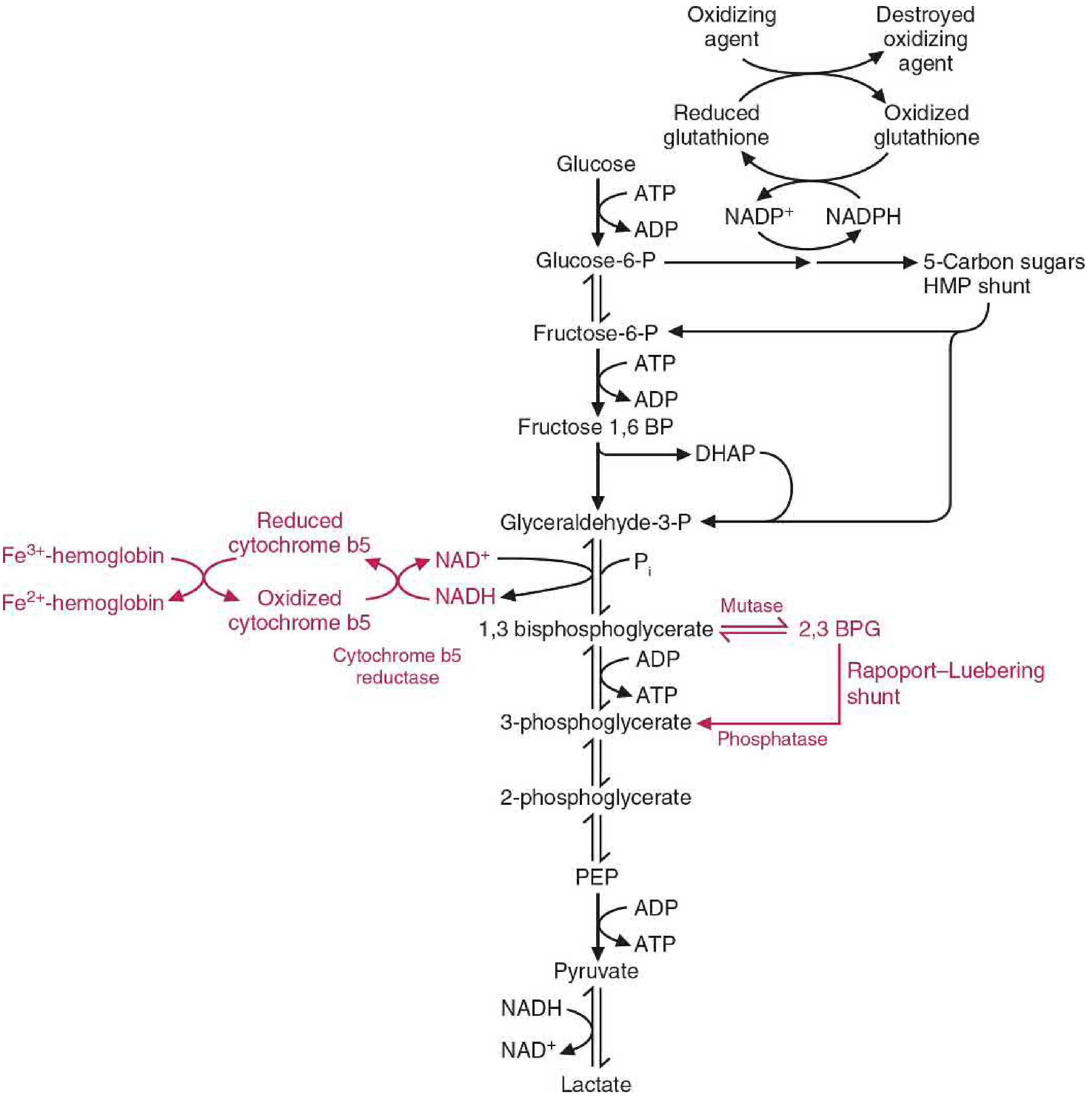
(Pathways unique to the RBC are shown in red; the Rapoport-Luebering shunt branches from 1,3-bisphosphoglycerate)
The ATP generated from glycolysis is used for:
- Ion transport across the cell membrane (Na⁺, K⁺, Ca²⁺)
- Phosphorylation of membrane proteins
- Priming reactions of glycolysis itself
The Rapoport-Luebering (RL) Cycle - Detail
What is it?
The Rapoport-Luebering shunt is a glycolytic bypass found exclusively in erythrocytes. In normal (non-RBC) glycolysis, 1,3-bisphosphoglycerate (1,3-BPG) is directly converted to 3-phosphoglycerate (3-PG) by phosphoglycerate kinase, generating 1 molecule of ATP per mole. In the RBC, this step can be bypassed through the RL shunt, which instead converts 1,3-BPG to 2,3-bisphosphoglycerate (2,3-BPG) - at the cost of that ATP.
Reactions of the RL Shunt
Step 1 - Mutase reaction:
1,3-Bisphosphoglycerate (1,3-BPG) → 2,3-Bisphosphoglycerate (2,3-BPG) Enzyme: 2,3-BPG mutase (BPGM, also called bisphosphoglycerate mutase)
Step 2 - Phosphatase reaction:
2,3-Bisphosphoglycerate (2,3-BPG) → 3-Phosphoglycerate (3-PG) + Inorganic phosphate (Pi) Enzyme: 2,3-BPG phosphatase (activity of the same BPGM enzyme)
3-Phosphoglycerate then re-enters the main glycolytic pathway and continues to pyruvate/lactate.
Diagram of the RL shunt and 2,3-BPG synthesis:
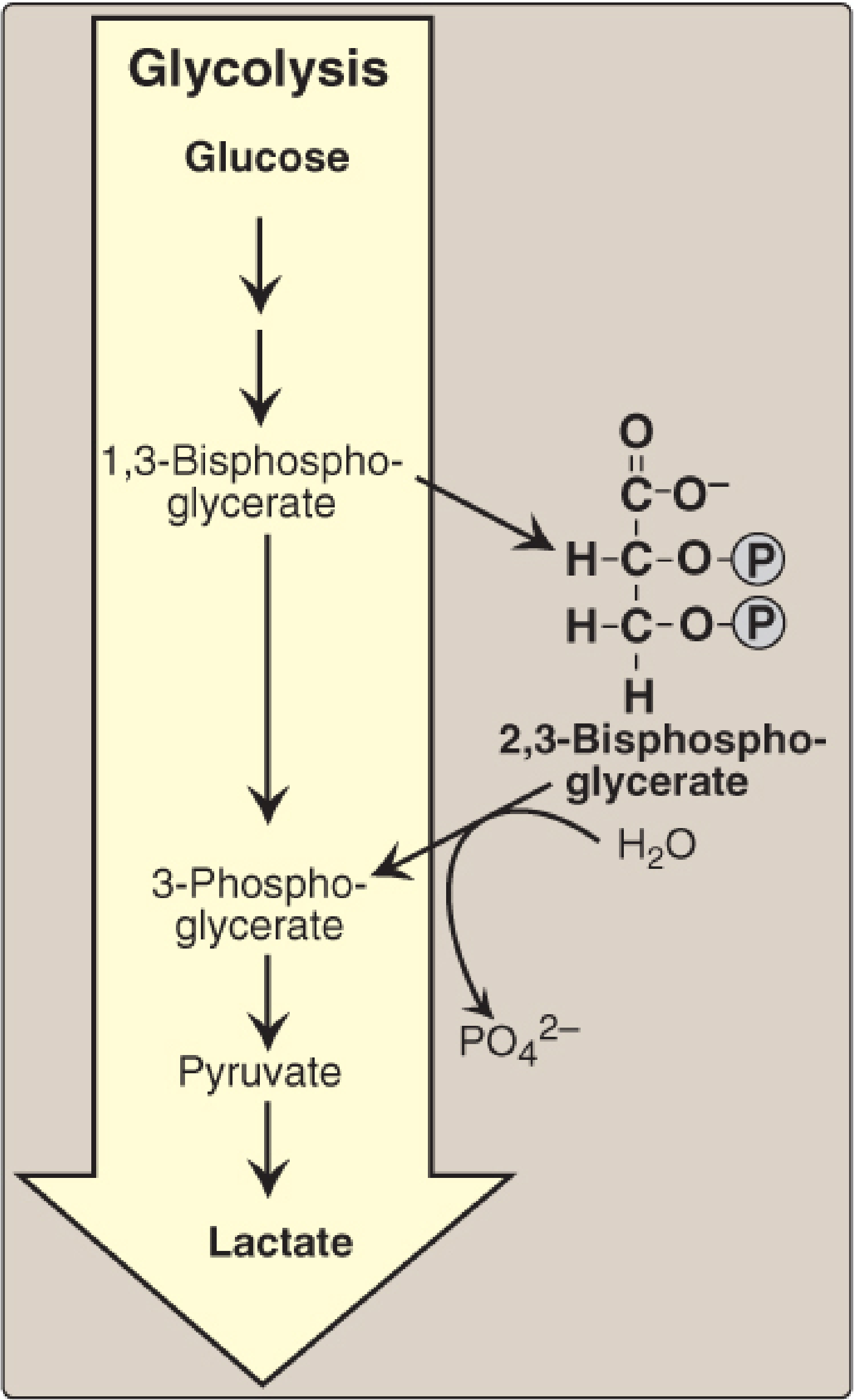
Enzyme - BPGM (Bisphosphoglycerate Mutase)
Both reactions of the RL shunt are catalyzed by a single multifunctional enzyme, BPGM:
- Has mutase activity: converts 1,3-BPG → 2,3-BPG
- Has phosphatase activity: converts 2,3-BPG → 3-PG
BPGM is a homodimer with 30 kDa subunits (258 amino acids). Its gene is located on chromosome 7q31-34 and is expressed only in erythroid tissue during late stages of differentiation. This is why 2,3-BPG is found in high concentrations (4-5 mM) only in RBCs, with trace amounts elsewhere.
Why Does the RBC Sacrifice ATP to Make 2,3-BPG?
The key trade-off: going through the RL shunt bypasses the phosphoglycerate kinase step, which means 1 ATP is lost per mole of glucose shunted. So the RL shunt produces no net ATP from the 1,3-BPG step. The RBC accepts this energetic cost because 2,3-BPG serves a critical physiological function.
Physiological Role of 2,3-BPG: Regulation of Oxygen Delivery
Mechanism
2,3-BPG is the most abundant organic phosphate in the RBC, present at approximately the same molar concentration as hemoglobin itself (~4-5 mM). Its primary function is to lower the oxygen affinity of hemoglobin, allowing efficient oxygen unloading to tissues.
Mechanism of action:
- 2,3-BPG binds to the central cavity formed by the two β-globin chains of deoxyhemoglobin (T-state/tense form)
- The binding pocket contains positively charged amino acids that form ionic bonds with the negatively charged phosphate groups of 2,3-BPG
- This binding stabilizes the T (deoxy) conformation of hemoglobin, increasing the energy required for the R-state (oxy) conformational change
- Consequently, hemoglobin releases oxygen more readily in tissues
Equilibrium:
HbO₂ + 2,3-BPG ⇌ Hb-2,3-BPG + O₂
Binding site of 2,3-BPG on deoxyhemoglobin:
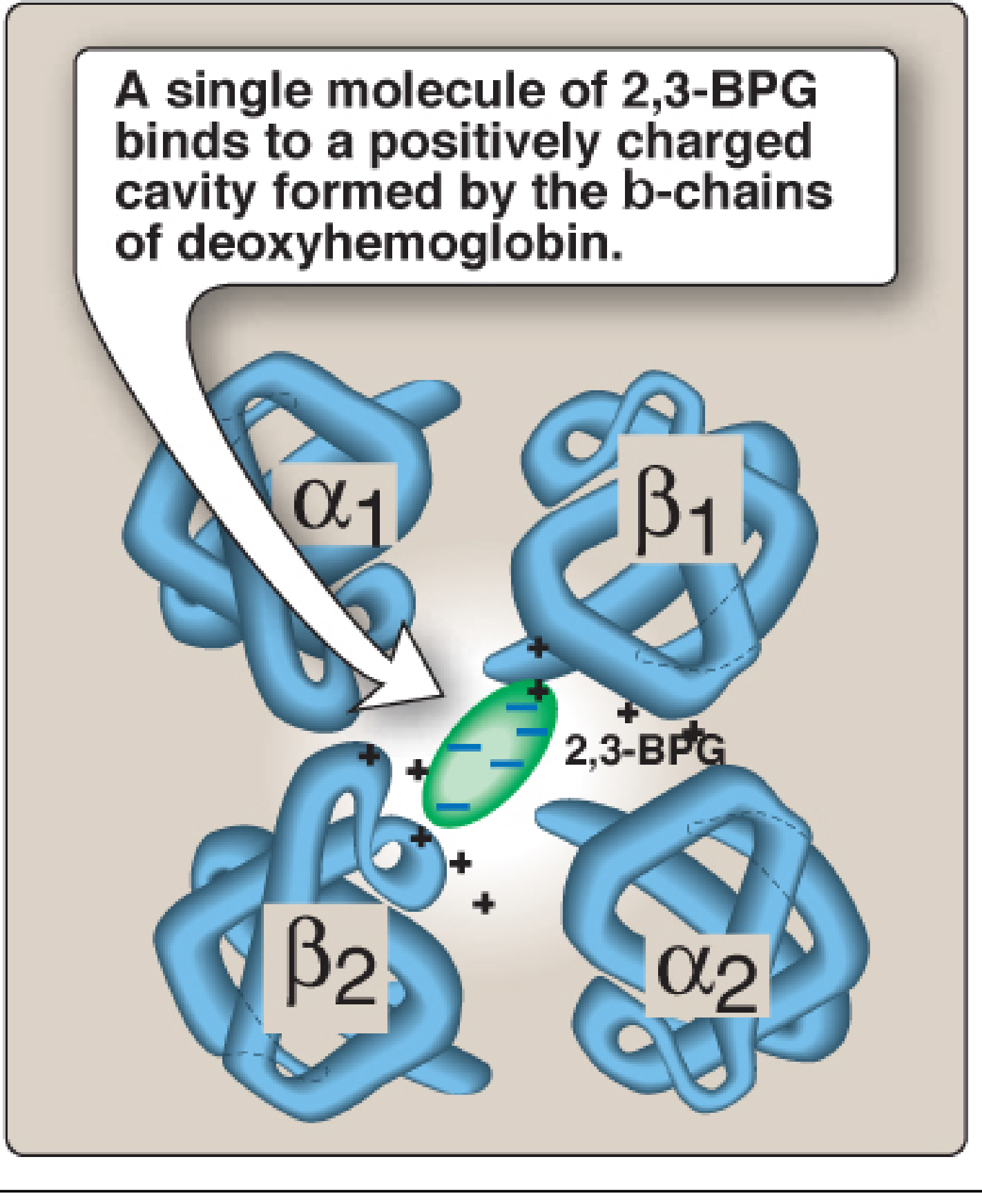
Key points about this binding:
- 2,3-BPG binds to deoxyhemoglobin but NOT to oxyhemoglobin
- When hemoglobin binds oxygen (becomes oxyhemoglobin), the central pocket narrows and 2,3-BPG is released
- This shifts the oxygen-dissociation curve to the right (decreased O₂ affinity = more O₂ released to tissues)
Conditions That Alter 2,3-BPG Levels
| Condition | 2,3-BPG Level | Effect on O₂ Delivery |
|---|---|---|
| High altitude / Hypoxia | Increased | More O₂ released to tissues (right shift) |
| Chronic anemia | Increased | Compensates for fewer RBCs |
| Stored blood (blood bank) | Decreased | Less O₂ delivered (left shift) - clinical concern |
| Pyruvate kinase deficiency | Increased | Compensates for hemolytic anemia |
| BPGM deficiency | Decreased | High O₂ affinity → erythrocytosis |
| HbF (fetal hemoglobin) | Binds 2,3-BPG weakly | Higher O₂ affinity than HbA |
Clinical Significance
1. Pyruvate Kinase (PK) Deficiency
PK deficiency blocks the conversion of phosphoenolpyruvate (PEP) to pyruvate, causing retrograde accumulation of glycolytic intermediates including 1,3-BPG. More 1,3-BPG is channeled into the RL shunt, resulting in 2-3 fold elevation of 2,3-BPG. This elevated 2,3-BPG partially compensates for the hemolytic anemia by facilitating better oxygen unloading in tissues. The ATP deficit from PK deficiency causes RBCs to become rigid and susceptible to destruction.
2. Stored Blood
During blood bank storage, 2,3-BPG levels in stored RBCs fall progressively (within 24-48 hours). This increases hemoglobin's oxygen affinity, meaning transfused blood may not deliver oxygen as effectively. 2,3-BPG levels are restored after transfusion within 24 hours as glycolysis resumes.
3. BPGM Deficiency (OMIM 222800)
A very rare autosomal recessive disorder. Absence of BPGM leads to reduced 2,3-BPG, so hemoglobin has very high oxygen affinity. This decreases tissue oxygenation, triggering compensatory erythrocytosis (increased RBC production). Only five families have been described in the literature.
4. Fetal Hemoglobin (HbF)
HbF has gamma (γ) chains instead of beta (β) chains. The γ-chains bind 2,3-BPG weakly because the key binding residue is different. Therefore, HbF has a higher oxygen affinity than adult HbA, which allows the fetus to extract oxygen from maternal blood across the placenta.
Summary of the Rapoport-Luebering Cycle
Glucose
↓ (Glycolysis - EMP)
1,3-Bisphosphoglycerate (1,3-BPG)
| |
| (Phosphoglycerate | (BPGM - Mutase activity)
| kinase - normal |
| glycolysis) ↓
| 2,3-Bisphosphoglycerate (2,3-BPG)
| |
↓ | (BPGM - Phosphatase activity)
3-Phosphoglycerate ←----+
↓
... → Pyruvate → Lactate
Effect: 2,3-BPG binds β-chains of deoxyhemoglobin
→ Stabilizes T-state → ↓O₂ affinity → More O₂ released to tissues
Key Points to Remember:
- The RL shunt is unique to RBCs
- It bypasses the ATP-generating phosphoglycerate kinase step (costs 1 ATP)
- Both reactions are catalyzed by one enzyme: BPGM (on chromosome 7q31-34)
- RBCs contain 4-5 mM 2,3-BPG (far more than any other cell type)
- 2,3-BPG binds only to deoxyhemoglobin (T-state), stabilizing it and shifting the O₂-dissociation curve to the right
- Increased 2,3-BPG is a physiological response to hypoxia and anemia
Sources: Basic Medical Biochemistry - A Clinical Approach, 6e; Lippincott's Illustrated Reviews: Biochemistry, 8e; Tietz Textbook of Laboratory Medicine, 7e
This is a shared conversation. Sign in to Orris to start your own chat.