I now have all the content and diagrams needed. Let me compile the comprehensive answer:
Smooth Muscle - Complete MBBS 1st Year Physiology Notes
Primary source: Guyton and Hall Textbook of Medical Physiology, 14th Edition; Costanzo Physiology, 7th Edition
1. Structure of Smooth Muscle Cells
Smooth muscle fibers are small, spindle-shaped cells - typically 1-5 micrometers in diameter and 20-500 micrometers in length - making them roughly 30 times narrower and hundreds of times shorter than skeletal muscle fibers.
Key structural features:
- No striations - because thick (myosin) and thin (actin) filaments are present but NOT organized into sarcomeres. This is why smooth muscle looks "smooth" under light microscopy.
- Single, centrally placed nucleus
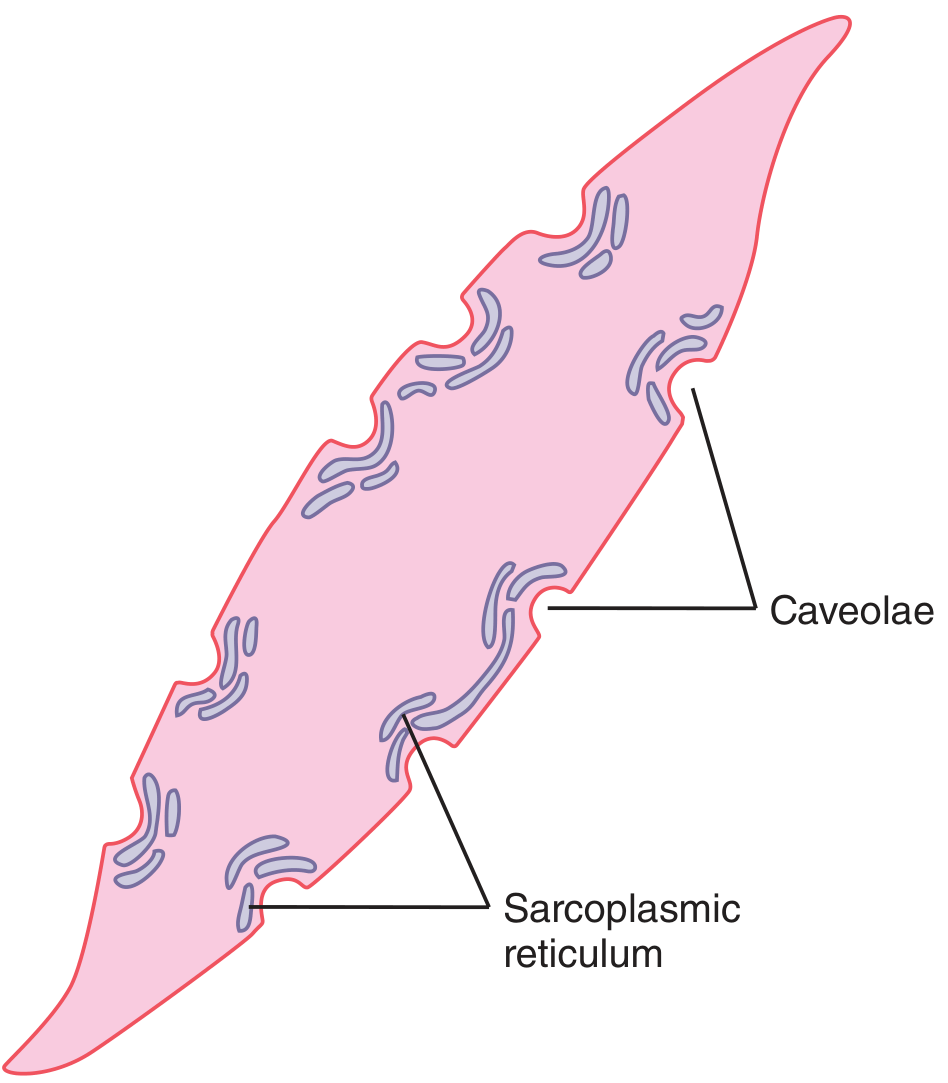
- Caveolae - small flask-shaped invaginations of the cell membrane that abut slightly developed sarcoplasmic tubules. These are the smooth muscle equivalent of the transverse tubule system in skeletal muscle. When an action potential enters the caveolae, it triggers Ca²⁺ release from the abutting sarcoplasmic reticulum.
- Sarcoplasmic reticulum - sparse and only slightly developed compared to skeletal muscle (see diagram below). Most Ca²⁺ therefore enters from the extracellular fluid rather than being released from intracellular stores.
- Dense bodies - scattered throughout the cytoplasm and attached to the cell membrane, equivalent to Z-lines in skeletal muscle. Actin filaments are anchored to these.
- Intermediate filaments (desmin) - form the cytoskeletal scaffolding that connects dense bodies.
Figure: A smooth muscle cell showing caveolae (membrane invaginations) and the sparse sarcoplasmic reticulum - Guyton & Hall
2. Types of Smooth Muscle
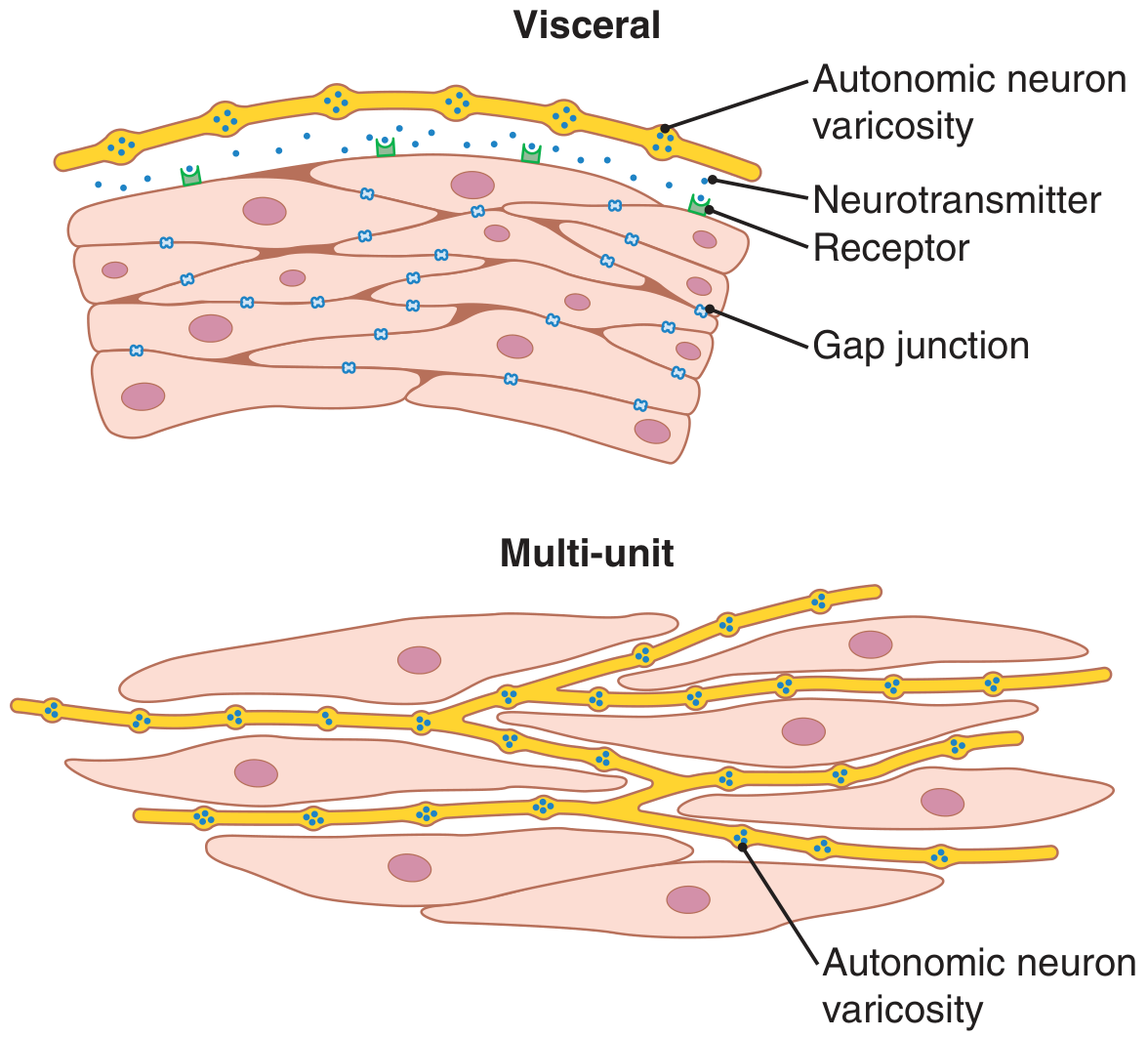
Smooth muscle is divided into two major types based on their electrical coupling and innervation:
Figure: Visceral (unitary) smooth muscle showing gap junctions enabling syncytial contraction (top) vs. multi-unit smooth muscle with individual innervation (bottom) - Guyton & Hall
Multi-Unit Smooth Muscle
- Composed of discrete, separate fibers, each operating independently
- Each fiber is often innervated by a single nerve ending, similar to skeletal muscle
- Outer surfaces are covered by a basement membrane-like substance (collagen + glycoprotein) that insulates fibers from each other
- Little or no gap junctions between cells - therefore no electrical coupling
- Each fiber acts as a separate motor unit
- Control is exerted mainly by nerve signals
- Examples: ciliary muscle of the eye, iris muscle, piloerector muscles of hair follicles, vas deferens
Unitary (Visceral / Syncytial) Smooth Muscle
- Hundreds to thousands of fibers arranged in sheets or bundles that contract together as a single unit ("unitary")
- Cell membranes are adherent at multiple points - force generated in one fiber transmits to the next
- Gap junctions connect adjacent cells - ions flow freely, so action potentials or depolarization currents spread from fiber to fiber
- This creates a functional syncytium (hence also called syncytial smooth muscle)
- Shows spontaneous pacemaker activity (slow waves) and can contract without direct nervous input
- Examples: walls of the gastrointestinal tract, bile ducts, ureters, uterus, urinary bladder, and many blood vessels
| Feature | Multi-Unit | Unitary (Visceral) |
|---|
| Cell coupling | None (no gap junctions) | Gap junctions - syncytium |
| Innervation | Each fiber individually | Only outer layer; spreads inward |
| Spontaneous activity | Absent | Present (pacemaker/slow waves) |
| Control | Primarily neural | Neural + hormonal + stretch |
| Examples | Iris, ciliary muscle, piloerector | GIT, uterus, bladder, ureters |
3. Membrane Potential and Action Potentials in Smooth Muscle
Resting Membrane Potential
The resting membrane potential of smooth muscle is approximately -50 to -60 millivolts - about 30 mV less negative than skeletal muscle (-90 mV). This lower resting potential means smooth muscle is closer to its firing threshold at baseline.
Action Potentials in Unitary Smooth Muscle
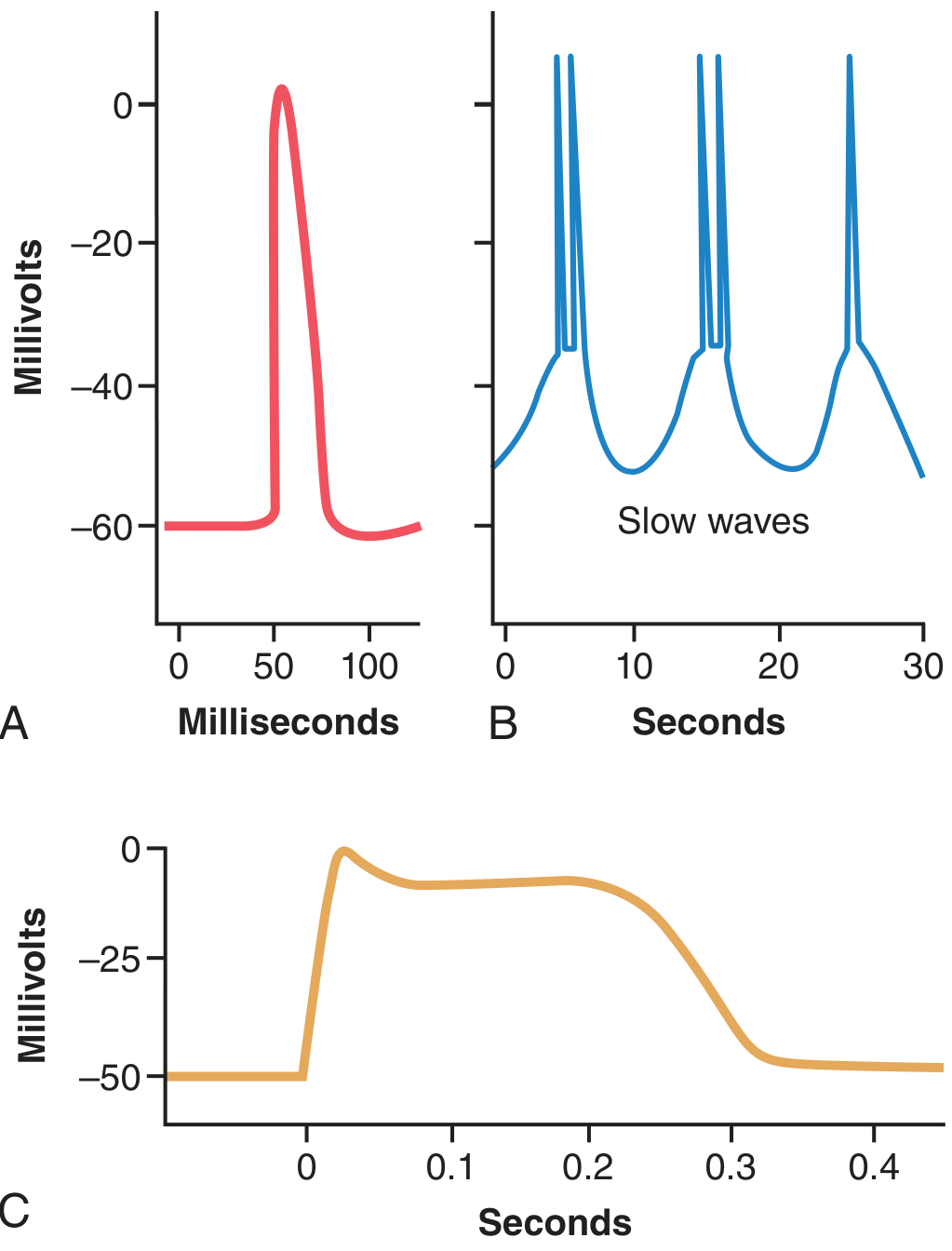
Action potentials occur readily in unitary smooth muscle but usually do NOT occur in most multi-unit smooth muscle (explained below). Visceral smooth muscle action potentials take two forms:
A. Spike Potentials
- Typical fast action potentials, similar in shape to skeletal muscle spikes
- Duration: 10 to 50 milliseconds
- Can be triggered by: electrical stimulation, hormones, neurotransmitters, stretch, or spontaneous pacemaker activity
B. Action Potentials with Plateaus
- The upstroke is similar to a spike potential, but repolarization is delayed for several hundred milliseconds to 1 second
- This prolonged plateau is responsible for the sustained contraction of some smooth muscle types
- Examples: uterus (especially during labour), certain vascular smooth muscle, ureter
Figure: (A) Typical spike action potential, (B) Repetitive spikes riding on slow waves (gut), (C) Plateau action potential (uterus) - Guyton & Hall
Why Ca²⁺ Channels Dominate (Not Na⁺)
This is the key physiological distinction from skeletal muscle and nerve:
- Smooth muscle cell membrane has many voltage-gated Ca²⁺ channels but few voltage-gated Na⁺ channels
- Therefore, it is the inward flow of Ca²⁺ (not Na⁺) that is mainly responsible for the upstroke of the action potential
- Ca²⁺ channels open much more slowly than Na⁺ channels and remain open longer - this explains the prolonged plateau of certain smooth muscle action potentials
- Ca²⁺ entry performs a dual function: it both generates the action potential AND directly triggers contraction via the calmodulin pathway
4. Physiological Basis of Action Potentials in Unitary Smooth Muscle: Slow Wave Potentials
This is a unique feature of unitary smooth muscle.
What are slow waves?
- Spontaneous, rhythmic oscillations of membrane potential that occur in unitary (visceral) smooth muscle
- They are not action potentials - they are not self-regenerative and do not spread like action potentials
- They are a local property of the smooth muscle fibers
Cause of slow waves (two hypotheses):
- Rhythmic waxing and waning of the Na⁺-K⁺ pump activity - when pump is active, more Na⁺ is expelled and the membrane hyperpolarizes; when pump slows, membrane depolarizes
- Rhythmic changes in conductance of ion channels
How slow waves trigger action potentials:
- When the slow wave depolarizes the membrane from -60 mV to approximately -35 mV (the threshold for action potential generation in visceral smooth muscle), a full action potential fires
- This action potential spreads through the syncytium via gap junctions and triggers a coordinated contraction
- The slow waves set the frequency of contractions in hollow visceral organs (e.g., gastric slow waves: ~3/min; small intestine: ~12/min)
- Hence, slow waves are called pacemaker waves
Stretch and spontaneous activity:
When visceral smooth muscle is stretched, the resting membrane potential becomes less negative (partial depolarization), which added to ongoing slow waves pushes the membrane to threshold more easily - generating spontaneous action potentials. This is the basis of the gut's automatic response to distension (propulsive peristalsis).
5. Depolarization of Multi-Unit Smooth Muscle Without Action Potentials
In multi-unit smooth muscle (iris, piloerector muscles):
- Nerve endings release neurotransmitters (ACh from parasympathetic fibers; norepinephrine from sympathetic fibers)
- These transmitters cause local depolarization of the cell membrane called the junctional potential
- Action potentials usually do NOT develop because:
- The individual fibers are too small to generate a self-propagating action potential
- In visceral smooth muscle, a critical mass of 30-40 fibers must depolarize simultaneously before a self-propagating action potential can be initiated. Multi-unit fibers, being individually small and isolated, cannot achieve this
- The junctional potential spreads electrotonically (passively, without regeneration) over the entire fiber
- This electrotonic spread is sufficient to cause contraction without requiring a full action potential
- This makes multi-unit smooth muscle contraction fine and graded, since each fiber responds individually and in proportion to the strength of neurotransmitter stimulation
6. Local Tissue Factors and Hormones Causing Contraction Without Action Potentials
Smooth muscle (especially vascular smooth muscle) can contract in response to many local chemical changes without any action potential being generated. These stimuli act directly on the contractile proteins or on Ca²⁺ channels through second messengers.
Local Tissue Factors
| Factor | Effect |
|---|
| Lack of oxygen | Causes relaxation (vasodilation) |
| Excess CO₂ | Relaxation |
| Decreased pH (acidosis) | Relaxation |
| Increased K⁺ | Depolarization and contraction |
| Increased Ca²⁺ | Contraction |
| Adenosine | Vasodilation (smooth muscle relaxation) |
| Stretch | Contraction (myogenic response in vascular smooth muscle) |
Hormones
Smooth muscle membranes contain a large variety of receptor proteins that respond to circulating hormones:
- Norepinephrine / epinephrine: Can cause contraction (via α₁-receptors, IP₃/DAG pathway raising intracellular Ca²⁺) or relaxation (via β₂-receptors, cAMP pathway)
- Acetylcholine: Contracts most visceral smooth muscle; relaxes some (via endothelium-derived NO in vessels)
- Oxytocin: Contracts uterine smooth muscle
- Angiotensin II: Contracts vascular smooth muscle
- Vasopressin (ADH): Contracts vascular smooth muscle
- Histamine: Contracts bronchial smooth muscle (causes bronchoconstriction in asthma)
- Serotonin (5-HT): Contracts GI smooth muscle
- Prostaglandins: Variable effects depending on type and receptor
Mechanism of hormone-induced contraction without action potential:
- Hormone binds its receptor → activates Gq protein → activates phospholipase C → generates IP₃ + DAG
- IP₃ releases Ca²⁺ from sarcoplasmic reticulum (intracellular stores)
- This rise in intracellular Ca²⁺ activates the calmodulin-MLCK pathway (see Section 7) and causes contraction WITHOUT the membrane ever reaching action potential threshold
7. Regulation of Smooth Muscle Contraction by Calcium Ions
This is the central mechanism of smooth muscle contraction. Unlike skeletal muscle (which uses the troponin system), smooth muscle uses the calmodulin-myosin light chain kinase (MLCK) system.
Step-by-step mechanism:
-
Ca²⁺ entry/release: Ca²⁺ enters from extracellular fluid via voltage-gated Ca²⁺ channels (opened during action potentials) AND is released from the sarcoplasmic reticulum (via IP₃ receptors or ryanodine receptors in response to hormones/neurotransmitters). The latent period for Ca²⁺ diffusion averages 200-300 ms (50x longer than skeletal muscle).
-
Calmodulin binding: The entering Ca²⁺ binds to calmodulin (CaM), a regulatory protein that is the functional analog of troponin C. Four Ca²⁺ ions bind per calmodulin molecule.
-
MLCK activation: The Ca²⁺-CaM complex activates myosin light chain kinase (MLCK), a phosphorylating enzyme.
-
Myosin phosphorylation: MLCK phosphorylates the regulatory light chain on each myosin head. When this light chain is NOT phosphorylated, the myosin head cannot interact with actin (cross-bridge cycling is impossible). When phosphorylated, the myosin head can bind to actin and undergo the full power-stroke cycle.
-
Cross-bridge cycling and contraction: The phosphorylated myosin heads repetitively attach to and detach from actin filaments, pulling them toward the center - shortening the cell and producing force. ADP and inorganic phosphate are released; ATP binds to allow detachment.
-
Relaxation: When intracellular Ca²⁺ falls, Ca²⁺ dissociates from calmodulin → MLCK becomes inactive → myosin light chain phosphatase (MLCP) dephosphorylates the regulatory light chain → cross-bridge cycling stops → the muscle relaxes.
Role of extracellular vs. intracellular Ca²⁺:
- The sarcoplasmic reticulum is only slightly developed in smooth muscle
- Therefore, the majority of activating Ca²⁺ comes from extracellular fluid (extracellular [Ca²⁺] = >10⁻³ M; intracellular [Ca²⁺] = <10⁻⁷ M - a 10,000-fold gradient)
- Removing Ca²⁺ from extracellular fluid will abolish smooth muscle contraction much more readily than in skeletal muscle
8. The Latch Mechanism
The latch mechanism is one of the most important and unique features of smooth muscle physiology. It explains how smooth muscle can maintain prolonged tonic contractions with minimal energy expenditure.
What is the Latch Mechanism?
When smooth muscle contracts, some myosin cross-bridges enter a special state where they remain attached to actin filaments for a prolonged period - they become "latched" or locked - even after the intracellular Ca²⁺ concentration has fallen. This is the latch state.
Physiological Basis
- Initially, elevated Ca²⁺ → MLCK phosphorylates myosin → active cross-bridge cycling (high energy use, fast cycling)
- As Ca²⁺ slowly falls, MLCP begins dephosphorylating the myosin light chains
- However, some dephosphorylated (unlocked) myosin heads remain attached to actin without cycling - they become "latch bridges"
- These latch bridges generate sustained tonic force but do NOT cycle repeatedly, so ATP consumption is dramatically reduced
- The latch state is eventually released by further Ca²⁺ reduction or by specific signaling
Physiological Significance
- Allows hours of sustained tonic contraction at very low metabolic cost (e.g., blood vessel tone, sphincter tone, bronchial tone)
- A blood vessel can maintain arterial pressure all day with negligible ATP consumption compared to what would be required for continuous active cross-bridge cycling
- This explains why smooth muscle is the ideal muscle for organs requiring sustained tension maintenance
9. Stress Relaxation and Reverse Stress Relaxation
These are unique mechanical properties of smooth muscle (especially visceral unitary smooth muscle) that are critical for the function of hollow organs.
Stress Relaxation (Receptive Relaxation)
Definition: When a hollow smooth-muscle organ is suddenly stretched (e.g., the bladder fills, or the stomach receives food), the wall tension initially rises sharply. However, over the next few seconds to minutes, the tension progressively falls back toward its original level - even though the volume has increased and the wall remains stretched.
Physiological mechanism:
- The myosin cross-bridges and structural proteins of smooth muscle can slowly rearrange and "slip" to new positions at the new length
- The latch bridges gradually release and re-form in the new elongated configuration
- The muscle effectively "resets" its resting length to the new stretched length
Physiological importance:
- Allows the stomach, bladder, and uterus to accommodate large volumes at low pressure
- A full bladder does not generate very high wall pressure until it is ready to void, preventing inadvertent micturition
- The stomach can expand from ~50 mL (empty) to >1 L (full meal) without generating excessively high pressures that would cause pain or back-pressure on the esophagus
Reverse Stress Relaxation
Definition: The opposite phenomenon. When a stretched hollow organ suddenly decreases in volume (e.g., after the bladder is partially emptied), the wall tension falls initially but then progressively rises back over subsequent seconds-minutes, tending to maintain a tonic contraction.
Physiological mechanism:
- When muscle shortens, cross-bridges and structural elements rearrange to the shorter configuration
- MLCK activity and Ca²⁺ signaling maintain partial activation
- The muscle "resets" again to the new shorter length and generates appropriate wall tension
Physiological importance:
- Helps the bladder maintain detrusor tone and continue expelling urine even as it empties
- Helps the uterus maintain coordinated contractions during labour
- Allows hollow organs to maintain tone across a wide range of volumes, adapting their wall tension proportionally
Quick Summary Table
| Topic | Key Point |
|---|
| Smooth muscle size | 1-5 μm diameter, 20-500 μm length; no striations |
| Multi-unit | Independent fibers, mainly neural control, no gap junctions |
| Unitary | Gap-junction syncytium, spontaneous slow waves, pacemaker activity |
| Resting potential | -50 to -60 mV (less negative than skeletal muscle) |
| Action potential carrier | Ca²⁺ (not Na⁺) via voltage-gated Ca²⁺ channels |
| Spike vs plateau AP | Spike: 10-50 ms; Plateau: up to 1000 ms (uterus, ureter) |
| Slow waves | Local oscillations; when reach -35 mV threshold → spike fired |
| Multi-unit contraction | Junctional potential (no AP); electrotonic spread sufficient |
| Ca²⁺ mechanism | Ca²⁺ → Calmodulin → MLCK → myosin phosphorylation → contraction |
| Latch mechanism | Dephosphorylated cross-bridges remain attached; tonic force at low energy cost |
| Stress relaxation | Stretch → initial tension rise → slow tension fall → accommodation of volume |
| Reverse stress relaxation | Shortening → initial tension fall → tension restoration → sustained contraction |
Sources: Guyton and Hall Textbook of Medical Physiology, 14th Ed., Chapter 8; Costanzo Physiology, 7th Ed., Chapter 1