Ketone body metabolism
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Reading File
Reading File
Loading Image
Loading Image
I now have comprehensive textbook content and diagrams. Let me compile a thorough answer on ketone body metabolism.
Ketone Body Metabolism
Ketone bodies are water-soluble lipid-derived fuels produced by the liver during states of low carbohydrate availability (fasting, starvation, prolonged exercise, uncontrolled diabetes). They serve as an alternative energy substrate for most tissues - including the brain - when glucose is scarce.
The Three Ketone Bodies
| Compound | Notes |
|---|---|
| Acetoacetate | Primary ketone body; organic acid; metabolically active |
| D-β-Hydroxybutyrate (3-hydroxybutyrate) | Most abundant in blood (~75%); technically a hydroxy acid, not a ketone chemically, but classified as a ketone body; metabolically active |
| Acetone | Formed by spontaneous decarboxylation of acetoacetate; volatile; exhaled via lungs; not metabolically utilized |
Normal blood ketone bodies: <3 mg/dL. The β-hydroxybutyrate:acetoacetate ratio is normally ~3:1 and rises with increasing NADH/NAD+ ratio.
1. Ketogenesis (Synthesis - Liver Mitochondria Only)
Ketogenesis occurs exclusively in hepatic mitochondria and is triggered when acetyl CoA from fatty acid β-oxidation floods the liver faster than the TCA cycle can handle it.
Why acetyl CoA accumulates:
- High fatty acid oxidation raises NADH, which inhibits the TCA cycle
- Elevated NADH shifts OAA → malate, depleting OAA for citrate synthase
- Without OAA, acetyl CoA cannot enter the TCA cycle efficiently
- Elevated acetyl CoA activates pyruvate carboxylase (driving gluconeogenesis) and inhibits pyruvate dehydrogenase
- The "overflow" acetyl CoA is diverted to ketone body synthesis
Steps of Ketogenesis:
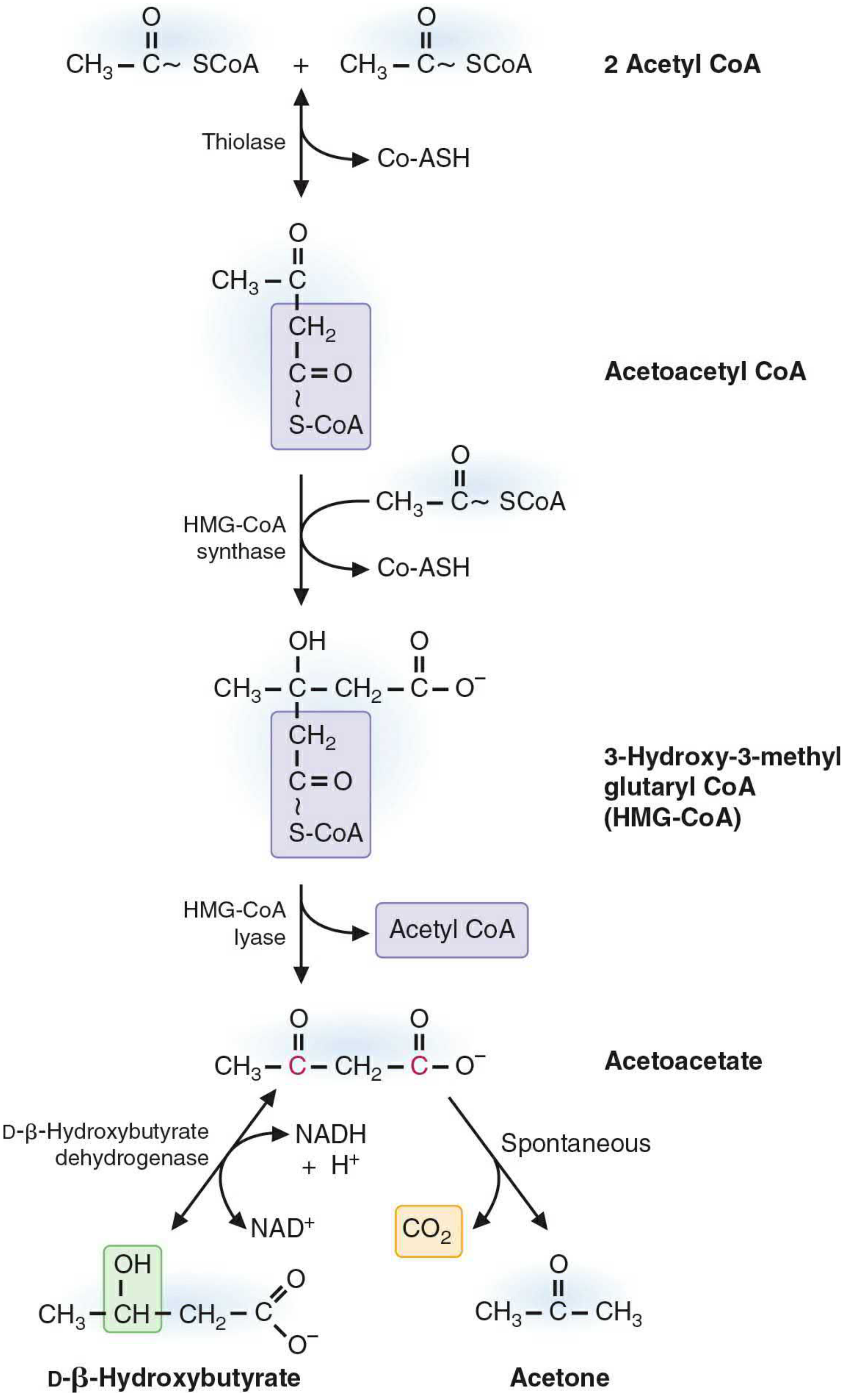
Step 1: Two acetyl CoA condense via thiolase (reversal of β-oxidation thiolase reaction) → Acetoacetyl CoA
Step 2 (Rate-limiting): Acetoacetyl CoA + Acetyl CoA → HMG-CoA (3-hydroxy-3-methylglutaryl CoA), catalyzed by mitochondrial HMG-CoA synthase
- This is the committed, rate-limiting step
- HMG-CoA synthase is present in significant quantities only in the liver
- Note: cytosolic HMG-CoA synthase exists separately for cholesterol synthesis
Step 3: HMG-CoA → Acetoacetate + Acetyl CoA, catalyzed by HMG-CoA lyase
Step 4a: Acetoacetate + NADH → D-β-Hydroxybutyrate, by β-hydroxybutyrate dehydrogenase (favored when NADH is high during fatty acid oxidation)
Step 4b: Acetoacetate → Acetone + CO₂ (spontaneous, non-enzymatic decarboxylation)
Because ketone bodies lack CoA, they freely cross the inner mitochondrial membrane and enter the bloodstream - Biochemistry, 8th ed, Lippincott Illustrated Reviews, p. 555-556
2. Ketone Body Transport
- Acetoacetate and β-hydroxybutyrate are water-soluble and travel freely in blood without a carrier protein (unlike long-chain fatty acids which need albumin)
- They are transported from liver to peripheral tissues via the bloodstream
3. Peripheral Utilization (Ketolysis)
Ketone bodies can be oxidized by skeletal muscle, cardiac muscle, renal cortex, intestinal mucosa, and - importantly during prolonged starvation - the brain.
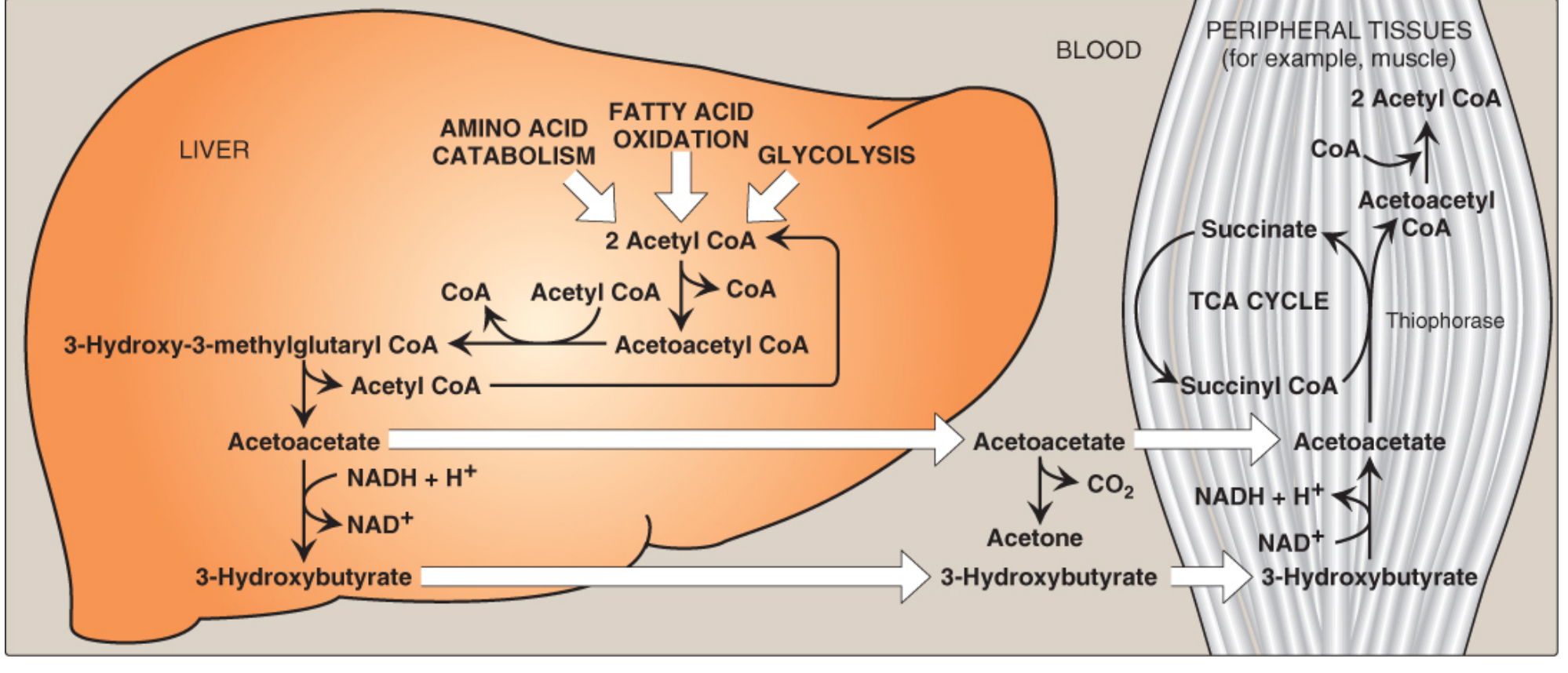
Steps of Ketolysis (in mitochondria of peripheral tissues):
Step 1: D-β-Hydroxybutyrate → Acetoacetate + NADH, by β-hydroxybutyrate dehydrogenase (reverse of ketogenesis step 4a)
Step 2: Acetoacetate + Succinyl CoA → Acetoacetyl CoA + Succinate, by succinyl CoA:acetoacetate CoA transferase (thiophorase)
- This is the activation step; succinyl CoA donates its CoA thioester bond
- In doing so, the GTP normally generated from succinyl CoA → succinate in the TCA cycle is foregone (costs 1 ATP equivalent per acetoacetate)
Step 3: Acetoacetyl CoA → 2 Acetyl CoA, by thiolase → enters TCA cycle → ATP
Energy Yield:
- Acetoacetate: ~19 ATP (net, accounting for 1 ATP equivalent activation cost)
- β-Hydroxybutyrate: ~21.5 ATP (one additional NADH from step 1)
Key restriction: The liver CANNOT use ketone bodies
- The liver lacks thiophorase (succinyl CoA:acetoacetate CoA transferase)
- This prevents futile cycling: the liver makes ketones but cannot consume them
- RBCs also cannot use ketone bodies (lack mitochondria)
4. Regulation of Ketogenesis
Ketogenesis is regulated primarily by substrate availability and hormonal signals:
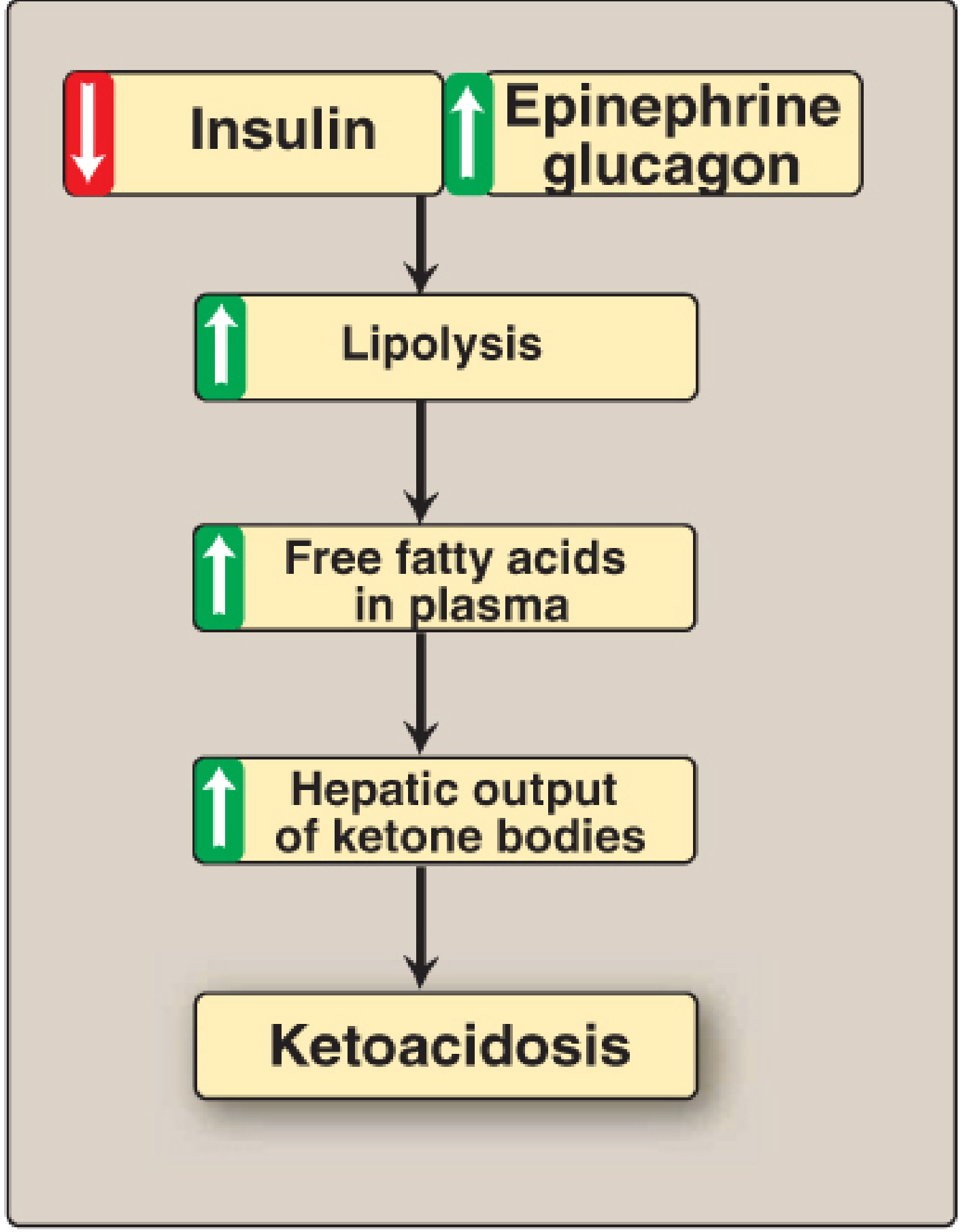
| Trigger | Effect on Ketogenesis |
|---|---|
| ↓ Insulin (fasting, T1D) | Activates HSL in adipose → lipolysis → FFA flood liver |
| ↑ Glucagon | Promotes hepatic fatty acid oxidation |
| ↑ Epinephrine | Stimulates lipolysis |
| ↑ Malonyl CoA (fed state) | Inhibits CPT-1 → blocks fatty acid entry into mitochondria → inhibits ketogenesis |
| ↑ NADH/NAD+ | Shifts OAA to malate → acetyl CoA diverted to ketone synthesis |
Malonyl CoA is a key regulatory molecule: it rises after meals (insulin-stimulated) and potently inhibits carnitine palmitoyltransferase-1 (CPT-1), the rate-limiting enzyme of fatty acid transport into mitochondria, thereby suppressing ketogenesis in the fed state.
5. Alternative Sources of Ketone Bodies
Ketone bodies can also arise from ketogenic amino acids (catabolized to acetyl CoA or acetoacetyl CoA):
- Purely ketogenic: Leucine, Lysine
- Both ketogenic and glucogenic: Isoleucine, Tryptophan, Phenylalanine, Tyrosine, Threonine
Additionally, acetoacetate can be activated to acetoacetyl CoA in the cytosol for cholesterol synthesis or cytosolic acetyl CoA production (e.g., for acetylcholine synthesis in neurons).
6. Physiological States
Fasting/Starvation
- After ~24-48 hrs of fasting, blood ketones rise to ~2 mM
- Brain shifts from ~0% to up to ~60-70% of energy from ketone bodies during prolonged starvation
- This spares muscle protein that would otherwise be catabolized for gluconeogenesis
- Children reach ketosis faster (higher brain:liver mass ratio, smaller glycogen stores)
Exercise
- Prolonged exercise promotes lipolysis; after exercise, sudden drop in muscle fatty acid use can transiently elevate hepatic ketone production
Diabetic Ketoacidosis (DKA)
In uncontrolled Type 1 diabetes: absent insulin → unrestrained lipolysis + increased glucagon → massive hepatic fatty acid influx → ketone bodies accumulate faster than tissues can use them
- Blood ketones can reach 90 mg/dL (vs <3 mg/dL normal)
- Urinary excretion up to 5,000 mg/24 hrs
- Each ketone body loses H+ at physiologic pH (pKa ~4) → metabolic acidosis
- Dehydration from glucosuria and ketonuria further concentrates H+
- Acetone on breath → fruity odor
- Biochemistry, 8th ed, Lippincott, p. 558
Alcoholic Ketoacidosis
- Ethanol oxidation via alcohol dehydrogenase raises hepatic NADH → depletes OAA → impairs TCA cycle → acetyl CoA diverted to ketone bodies
7. Clinical Applications of Ketogenic Diets
- Refractory epilepsy: High-fat, 3:1 lipid:carbohydrate ratio diet reduces seizure frequency (mechanism unclear)
- Pyruvate dehydrogenase deficiency: Ketone bodies bypass the block by providing acetyl CoA to the TCA cycle and cytosolic acetyl CoA for acetylcholine synthesis
- Medium-chain triglycerides (MCTs) induce ketosis more effectively than long-chain fats
- Basic Medical Biochemistry, p. 1760
Summary Pathway
Fatty acids (from adipose lipolysis)
↓ β-oxidation (liver mitochondria)
Acetyl CoA (excess)
↓ Thiolase
Acetoacetyl CoA
↓ HMG-CoA synthase [RATE-LIMITING]
HMG-CoA
↓ HMG-CoA lyase
Acetoacetate ──→ Acetone (exhaled; spontaneous)
↓ β-OH-butyrate dehydrogenase (NADH)
β-Hydroxybutyrate
↓↓ (blood transport to periphery)
β-Hydroxybutyrate → Acetoacetate → Acetoacetyl CoA (thiophorase)
↓ Thiolase
2 Acetyl CoA → TCA → ATP
Sources: Biochemistry, 8th ed - Lippincott Illustrated Reviews, pp. 555-559; Basic Medical Biochemistry, A Clinical Approach, 6e, pp. 1684-1773; Mulholland and Greenfield's Surgery, 7e, pp. 382-395
This is a shared conversation. Sign in to Orris to start your own chat.