Translation in eukaryotic cells
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
I have comprehensive information from the textbooks. Now I'll compose a thorough, well-structured answer.
Translation in Eukaryotic Cells
Translation is the process by which the nucleotide sequence of an mRNA is decoded into the amino acid sequence of a protein. In eukaryotes, translation occurs in the cytoplasm on 80S ribosomes and is divided into three main stages: initiation, elongation, and termination, followed by post-translational modifications.
Components Required
Before translation begins, the following must be present:
- 80S ribosomes (40S small subunit + 60S large subunit), each containing three tRNA-binding sites: A (aminoacyl), P (peptidyl), and E (exit)
- mRNA (monocistronic in eukaryotes - one coding region per mRNA, unlike prokaryotes)
- Aminoacyl-tRNAs - tRNAs charged with their specific amino acids by aminoacyl-tRNA synthetases
- Eukaryotic initiation factors (eIFs), elongation factors (eEFs), and release factors (eRF)
- GTP and ATP as energy sources
The Genetic Code & tRNA
Each codon is a three-nucleotide sequence on mRNA read 5'→3'. Of 64 possible codons, 61 specify amino acids and 3 are stop codons (UAA, UAG, UGA). The code is degenerate (multiple codons can specify the same amino acid).
tRNA links codons to amino acids. The anticodon region (7 nucleotides) base-pairs with the mRNA codon in an antiparallel fashion. The wobble hypothesis explains how the 5' nucleotide of the anticodon can form non-Watson-Crick base pairs with the 3' base of the codon, allowing a single tRNA species to recognize more than one codon.
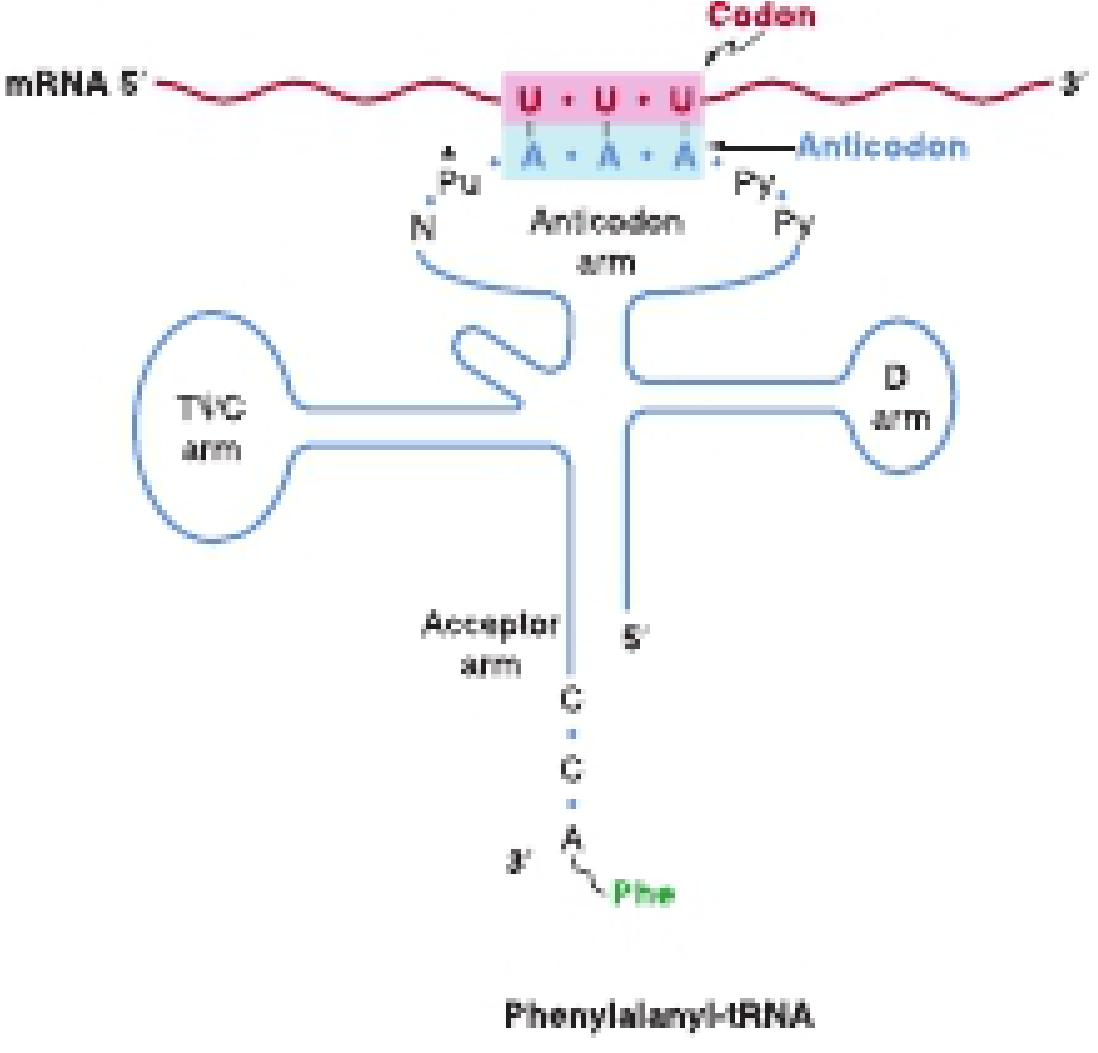
Aminoacyl-tRNA synthetases catalyze tRNA charging in a two-step reaction: (1) amino acid activation with ATP to form aminoacyl-AMP-enzyme complex, then (2) transfer of the amino acid to the 3'-OH of the tRNA. The error rate is extremely low (< 1 in 10^6 events). - Harper's Illustrated Biochemistry, 32nd Ed.
Stage 1: Initiation
Initiation assembles the full ribosome on the start codon. It involves at least 10 eukaryotic initiation factors (eIFs) and requires both GTP and ATP.
Step 1 - Ribosomal Dissociation: The 80S ribosome dissociates into 40S and 60S subunits at the end of each translation cycle. eIF-3, eIF-1, and eIF-1A bind the 40S subunit and prevent premature reassociation with the 60S subunit. - Harper's Illustrated Biochemistry, 32nd Ed.
Step 2 - 43S Preinitiation Complex: eIF-2 binds GTP, then recruits the initiator methionyl-tRNAi (Met-tRNAi) - a special tRNA used only at the start codon AUG (distinct from the tRNA used at internal AUG codons). This ternary complex (eIF-2•GTP•Met-tRNAi) joins the 40S subunit to form the 43S preinitiation complex, stabilized by eIF-3, eIF-1A, and eIF-5.
Step 3 - mRNA Binding and Scanning (48S complex): Unlike prokaryotes, eukaryotes lack a Shine-Dalgarno sequence. Instead, the small subunit (aided by eIF-4 family proteins) binds near the 5' cap structure (m7G cap) of the mRNA and scans 5'→3' until it encounters the first AUG codon. This scanning requires ATP hydrolysis. Interactions between cap-binding eIF-4 proteins and poly-A tail-binding proteins cause mRNA circularization, likely preventing use of incompletely processed mRNA. - Biochemistry, Lippincott 8th Ed.
Cap-independent initiation can also occur via binding of the 40S subunit to an Internal Ribosome Entry Site (IRES) near the start codon.
Step 4 - 80S Initiation Complex: Once the AUG is located, the 60S subunit joins the 48S complex (aided by eIF-5 and eIF-5B with GTP hydrolysis) to form the 80S initiation complex. The initiator Met-tRNAi occupies the P site (not the A site, unlike all subsequent aminoacyl-tRNAs). The A site is now empty and ready to accept the next aminoacyl-tRNA.
Key eukaryotic vs. prokaryotic differences: Eukaryotes use methionine as the initiating amino acid (prokaryotes use N-formylmethionine). The initiating codon is AUG in both.
Stage 2: Elongation
Elongation adds amino acids one at a time to the carboxyl end of the growing peptide. Each cycle has three steps:
1. Decoding (Aminoacyl-tRNA delivery to the A site):
The aminoacyl-tRNA matching the next codon in the A site is delivered by eEF-1α•GTP (equivalent to prokaryotic EF-Tu•GTP). When the correct anticodon base-pairs with the mRNA codon, GTP is hydrolyzed, eEF-1α•GDP is released, and the aminoacyl-tRNA is accommodated into the A site. The exchange factor eEF-1γ (EF-Ts analog) regenerates eEF-1α•GTP.
2. Peptide Bond Formation (Transpeptidation):
Peptidyl transferase - a ribozyme activity intrinsic to the 28S rRNA of the 60S subunit - catalyzes peptide bond formation between the α-carboxyl group of the amino acid in the P site and the α-amino group of the aminoacyl-tRNA in the A site. The growing peptide chain is transferred from the P-site tRNA to the A-site tRNA (transpeptidation), leaving an uncharged tRNA in the P site.
3. Translocation:
The ribosome moves exactly 3 nucleotides (one codon) toward the 3' end of the mRNA, facilitated by eEF-2•GTP (equivalent to prokaryotic EF-G) with GTP hydrolysis. This moves:
- The uncharged tRNA from P site → E site (then released)
- The peptidyl-tRNA from A site → P site
- The next codon into the A site
The cycle repeats until a stop codon enters the A site. Because mRNAs are long, many ribosomes can simultaneously translate a single mRNA, forming a polysome (polyribosome). - Biochemistry, Lippincott 8th Ed.
Stage 3: Termination
When one of the three stop codons (UAA, UAG, or UGA) moves into the A site, no aminoacyl-tRNA recognizes it. Instead:
- eRF-1 (a single eukaryotic release factor) recognizes all three stop codons
- eRF-1 triggers hydrolysis of the bond between the completed polypeptide and the P-site tRNA, releasing the protein
- eRF-3•GTP (analogous to prokaryotic RF-3) functions with eRF-1 and assists in release of eRF-1 upon GTP hydrolysis
- The ribosomal subunits, mRNA, and tRNA are released and recycled (requiring eRF and ATP hydrolysis in eukaryotes)
In prokaryotes, two release factors (RF-1 and RF-2) are needed; RF-1 recognizes UAA and UAG, RF-2 recognizes UGA and UAA.
Regulation of Translation
eIF-2 Phosphorylation (Key Control Point)
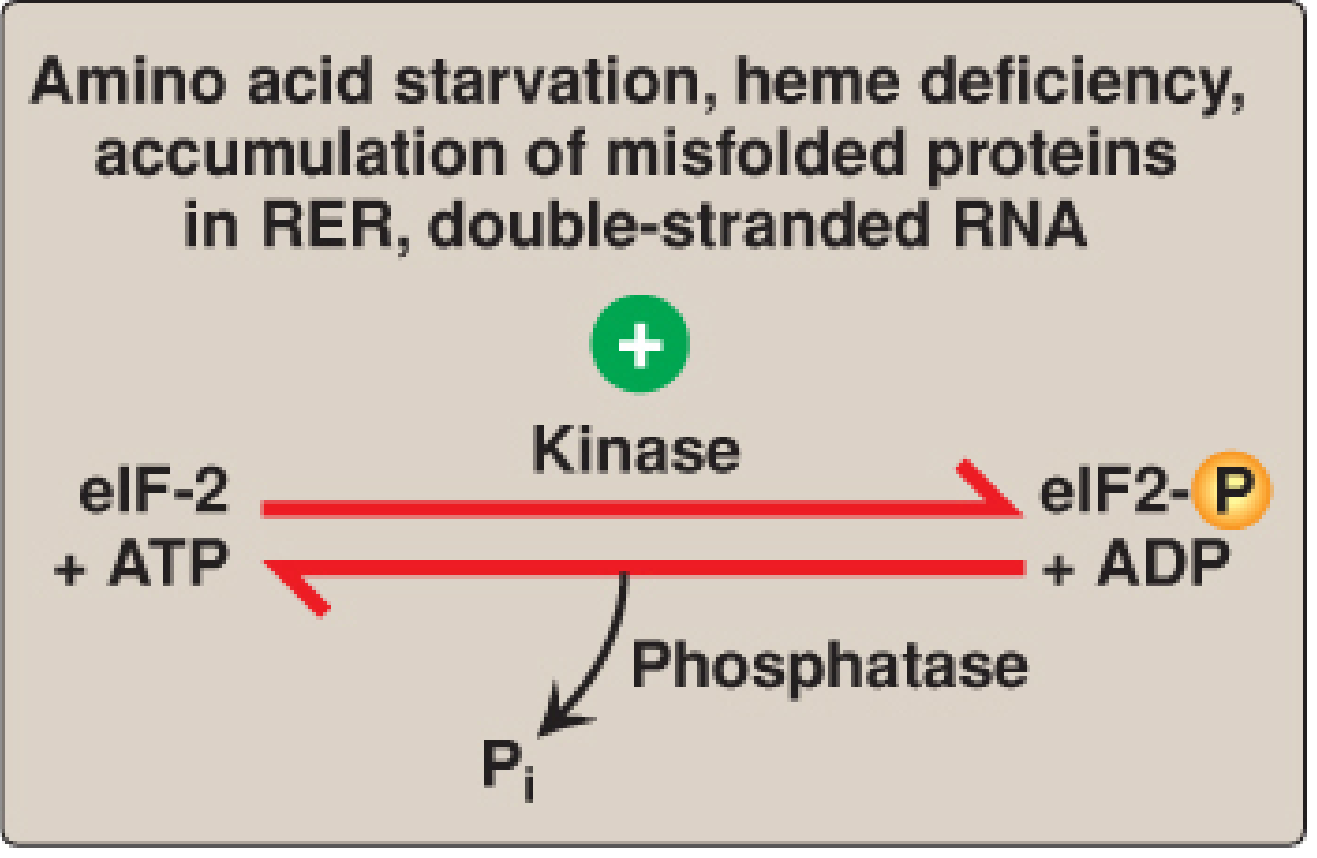
Under cellular stress conditions (amino acid starvation, heme deficiency, misfolded proteins in RER, viral infection with double-stranded RNA), specific kinases phosphorylate eIF-2α at Serine 51:
| Kinase | Trigger |
|---|---|
| HCR | Heme deficiency |
| PKR | Viral dsRNA |
| PERK | ER stress / misfolded proteins |
| GCN2 | Amino acid / glucose starvation |
Phosphorylated eIF-2α binds tightly to eIF-2B (the GEF that recycles GDP→GTP on eIF-2), inactivating it. This prevents formation of the 43S preinitiation complex and globally shuts down protein synthesis. This is a protective mechanism - PKR in particular provides antiviral defense by halting viral protein synthesis.
mTOR and eIF-4E-BP1
The mTOR pathway also regulates translation initiation. When active, mTOR phosphorylates 4E-BP1 (eIF-4E-binding protein), causing it to release eIF-4E. Free eIF-4E can then bind the 5' cap and initiate translation. When mTOR is inactive, 4E-BP1 sequesters eIF-4E, blocking translation of specific mRNAs.
Post-Translational Processing
After translation, proteins may undergo:
- Cleavage - removal of signal sequences or propeptides
- Covalent modifications - phosphorylation, glycosylation, acetylation, hydroxylation, etc.
- Folding - assisted by chaperone proteins
- Targeting - signal sequences direct proteins to organelles or for secretion
Summary Comparison: Eukaryotes vs. Prokaryotes
| Feature | Eukaryotes | Prokaryotes |
|---|---|---|
| Ribosome | 80S (40S + 60S) | 70S (30S + 50S) |
| mRNA type | Monocistronic | Polycistronic |
| Start amino acid | Methionine | N-formylmethionine |
| AUG recognition | 5' cap + scanning | Shine-Dalgarno sequence |
| Initiation factors | ≥10 eIFs (complex) | 3 IFs (simpler) |
| Elongation factors | eEF-1α, eEF-1γ, eEF-2 | EF-Tu, EF-Ts, EF-G |
| Release factors | eRF-1 (recognizes all 3 stops) | RF-1, RF-2 (separate) |
| Coupling of transcription/translation | No (nuclear membrane separates them) | Yes (simultaneous) |
| Energy for initiation | GTP + ATP | GTP |
Sources: Biochemistry, Lippincott Illustrated Reviews 8th Ed., pp. 1254-1270; Harper's Illustrated Biochemistry 32nd Ed., pp. 418-440
This is a shared conversation. Sign in to Orris to start your own chat.