Theories of olfaction in detail for 20 marks with diagram and flow charts from Scott Brown, Cummings, stell and marans, Zakir Hussain, dhingra, hazarika and related articles with recent advances according to rguhs university
Reading File
Finding Sources
Finding Sources
Searching PubMed
"olfaction" AND "theories" OR "olfactory transduction"
Reading File
Reading File
Reading File
Reading File
Searching PubMed
"olfactory receptor" AND "signal transduction" AND "mechanism"
Reading File
Reading File
Finding Sources
Finding Sources
Searching PubMed
"olfactory" AND "receptor neurons" AND "transduction" AND "recent advances"
Loading Image
Loading Image
Reading File
Reading File
Reading File
Reading File
Finding Sources
Searching PubMed
"olfactory bulb" AND "odor coding" AND "glomeruli"
Reading File
Reading File
Reading File
Searching the Web
theories of olfaction stereochemical vibration enzyme induction Amoore Dyson Beets RGUHS ENT
Loading Image
Loading Image
Reading File
Reading File
Loading Image
Loading Image
Searching PubMed
"olfactory" AND "Nobel Prize" AND "Buck Axel" OR "odorant receptor" AND "advances"
Searching the Web
theories of olfaction Dhingra Hazarika textbook ENT smell mechanism stereochemical vibration
I now have comprehensive content from all the textbooks and the web. Let me compile the complete, detailed answer.
THEORIES OF OLFACTION
A Comprehensive 20-Mark Answer for RGUHS University
Compiled from Scott-Brown's Otorhinolaryngology (Vol. 1 & 2), Cummings Otolaryngology Head & Neck Surgery, Costanzo Physiology, Guyton & Hall, Ganong's Review, Neuroscience: Exploring the Brain, Medical Physiology (Boron & Boulpaep), Bradley & Daroff's Neurology, and related recent literature.
INTRODUCTION
Olfaction (the sense of smell) is the detection of volatile chemical stimuli by specialized neuroepithelium in the nasal cavity and their transduction into neural signals interpreted by the brain. As noted in Cummings Otolaryngology, olfaction serves as a warning system for environmental hazards, facilitates social communication, and guides food intake. Impairment significantly reduces quality of life and is associated with depression.
The fundamental question in olfactory science - How do we recognize and discriminate odors? - has generated multiple competing theories over two centuries.
PART I: ANATOMY OF THE OLFACTORY SYSTEM
A. Olfactory Neuroepithelium (OE)
The OE is a pseudostratified columnar epithelium lining the olfactory cleft, spanning approximately 2-5 cm² in young adults. It contains three main cell types (Cummings, Ch. 36):
| Cell Type | Location | Function |
|---|---|---|
| Olfactory Receptor Neurons (ORN) | Mid-layer, bipolar neurons | Odorant binding, transduction, signal transmission |
| Supporting (Sustenacular) Cells | Columnar cells flanking ORN | Metabolic support, electrical isolation, mucus secretion |
| Basal Cells | Base of epithelium | Stem cells - generate new ORN (adult neurogenesis) |
| Bowman Glands | Lamina propria | Secrete mucus overlying OE |
Diagram 1: Olfactory Neuroepithelium (from Cummings / Costanzo)
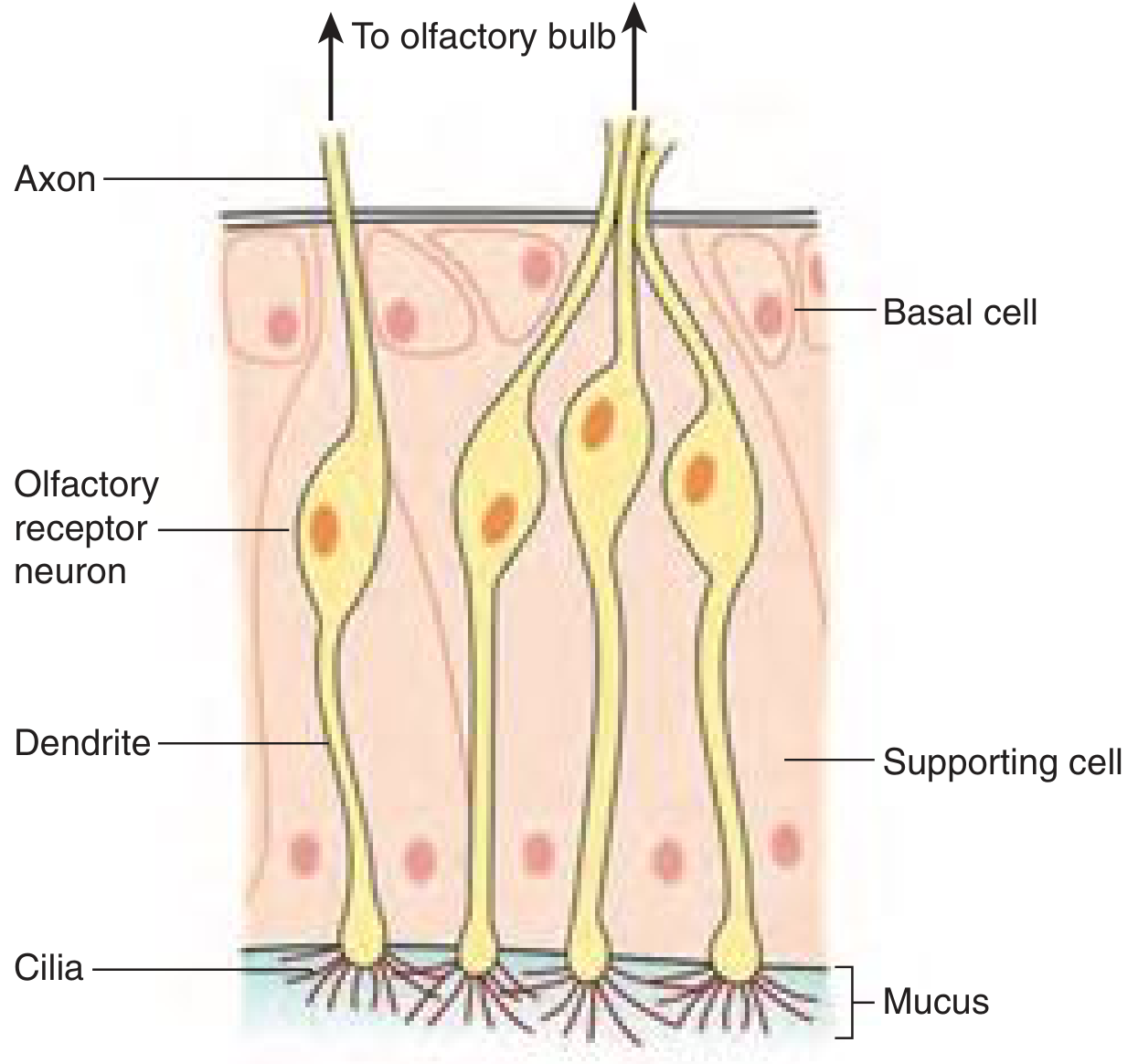
Fig. 1 - The olfactory neuroepithelium showing the olfactory receptor neuron with luminal dendrite and cilia, and basal axon extending toward the olfactory bulb. (Cummings Otolaryngology, Fig. 36.1)
Key structural features of ORN:
- Bipolar neurons: 6-30 million in young adults
- Single dendrite terminates in olfactory knob bearing 4-25 non-motile cilia (0.3 µm diameter, up to 200 µm long)
- Cilia project into mucous layer and contain olfactory receptors (ORs)
- Single unmyelinated axon passes through cribriform plate foramina to synapse in the ipsilateral olfactory bulb (OB)
- Adult neurogenesis: ORN are replaced from basal cells every 4-6 weeks (animals); rate in humans unknown
B. Olfactory Bulb and Higher Pathways
Diagram 2: Olfactory Bulb Structure and Connections
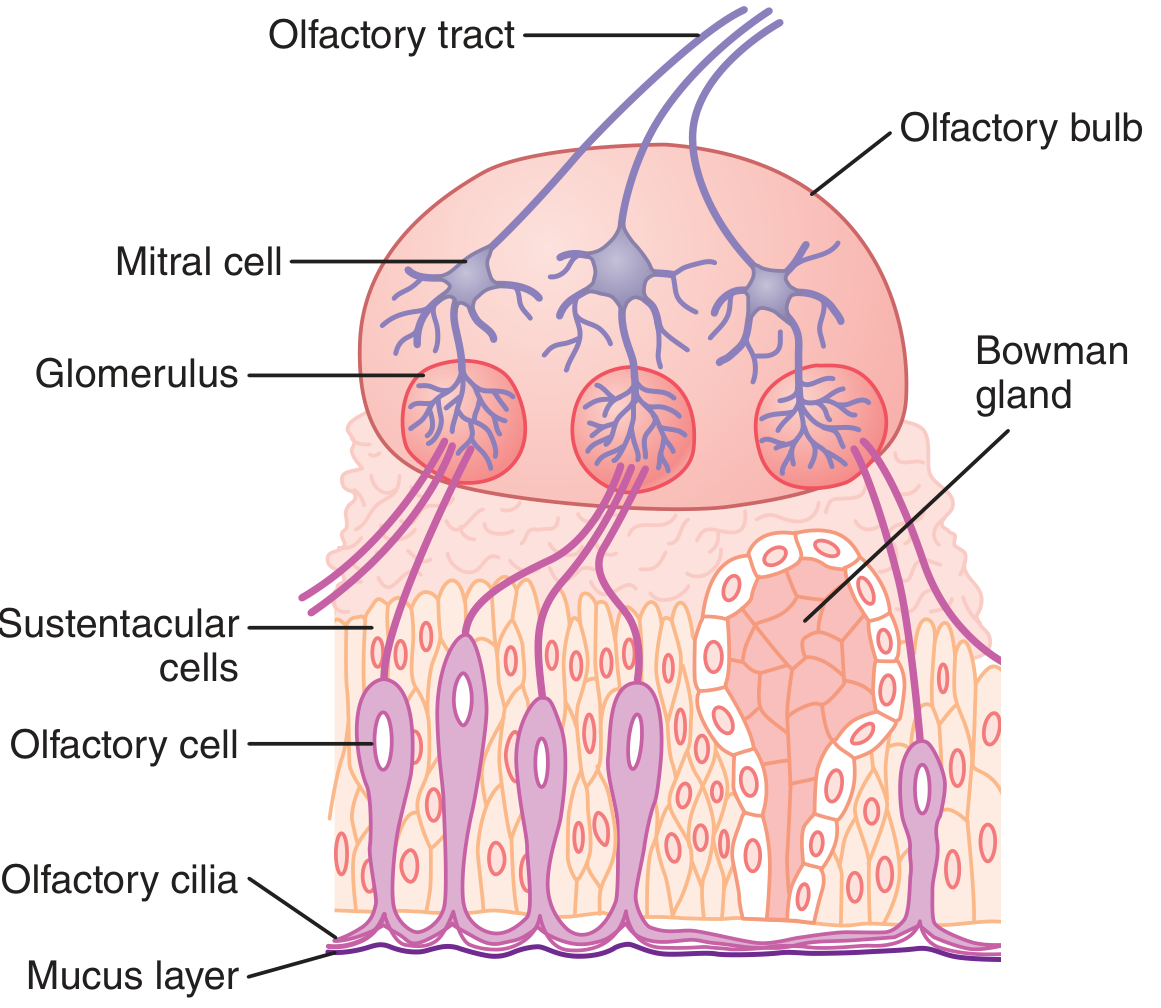
Fig. 2 - Organization of the olfactory membrane and olfactory bulb with connections to the olfactory tract. (Guyton & Hall, Fig. 54.3)
Diagram 3: Anatomical pathway from nasal cavity to olfactory bulb
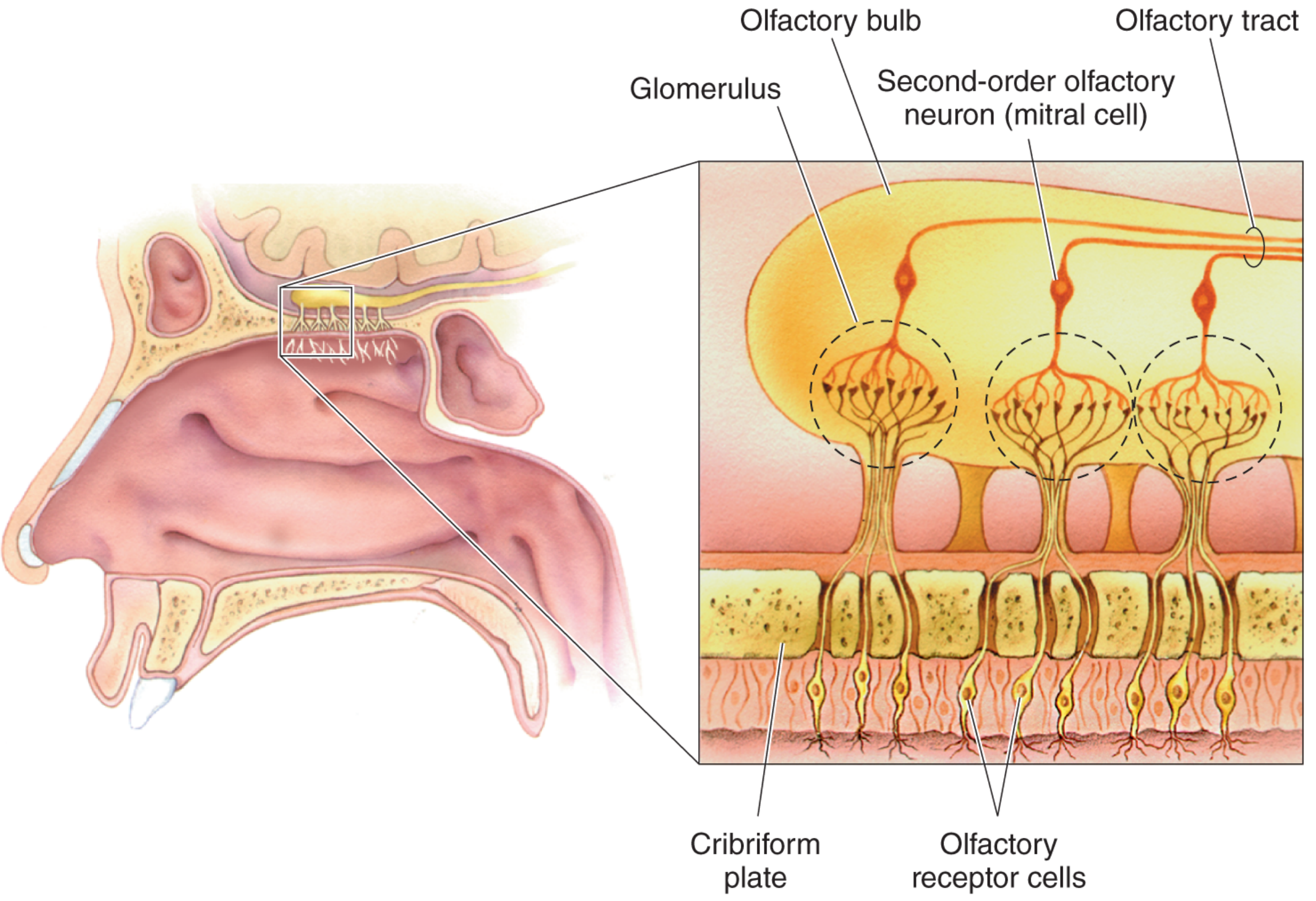
Fig. 3 - Location and structure of the olfactory bulb. ORN axons penetrate the cribriform plate, enter the olfactory bulb, and synapse on mitral cells within glomeruli. (Neuroscience: Exploring the Brain, Fig. 8.17)
Olfactory Bulb (OB) circuit (Ganong's Review, p. 188):
- ORN axons synapse on mitral cells and tufted cells within glomeruli (spherical tangles of ~25,000 ORN axons converging onto ~100 mitral cells)
- Periglomerular cells: inhibitory interneurons connecting glomeruli - sharpen olfactory signals
- Granule cells: no axons; make reciprocal synapses with mitral/tufted cell lateral dendrites; mediate lateral inhibition via GABA
- Each olfactory sensory neuron expresses only one of the ~400 functional olfactory receptor genes; projects to only 1-2 glomeruli - creating a unique 2D map in the OB
Higher olfactory projections (Ganong's Review):
Mitral/tufted cell axons travel via the lateral olfactory stria to five regions:
- Anterior olfactory nucleus
- Olfactory tubercle
- Piriform cortex (primary olfactory cortex)
- Amygdala (emotional/memory component)
- Entorhinal cortex
From these, information travels to the orbitofrontal cortex (via thalamus) for conscious odor discrimination.
Note: Unlike other sensory systems, olfaction does NOT relay through the thalamus as a primary route - it projects directly to cortex, which explains its strong emotional and memory associations.
PART II: OLFACTORY TRANSDUCTION - THE MOLECULAR MECHANISM
This is the modern "accepted theory" of olfaction - the G-Protein Coupled Receptor (GPCR) / cAMP Cascade Theory.
Steps in Olfactory Transduction
Flow Chart 1: Olfactory Signal Transduction (Costanzo Physiology, Fig. 3.27)
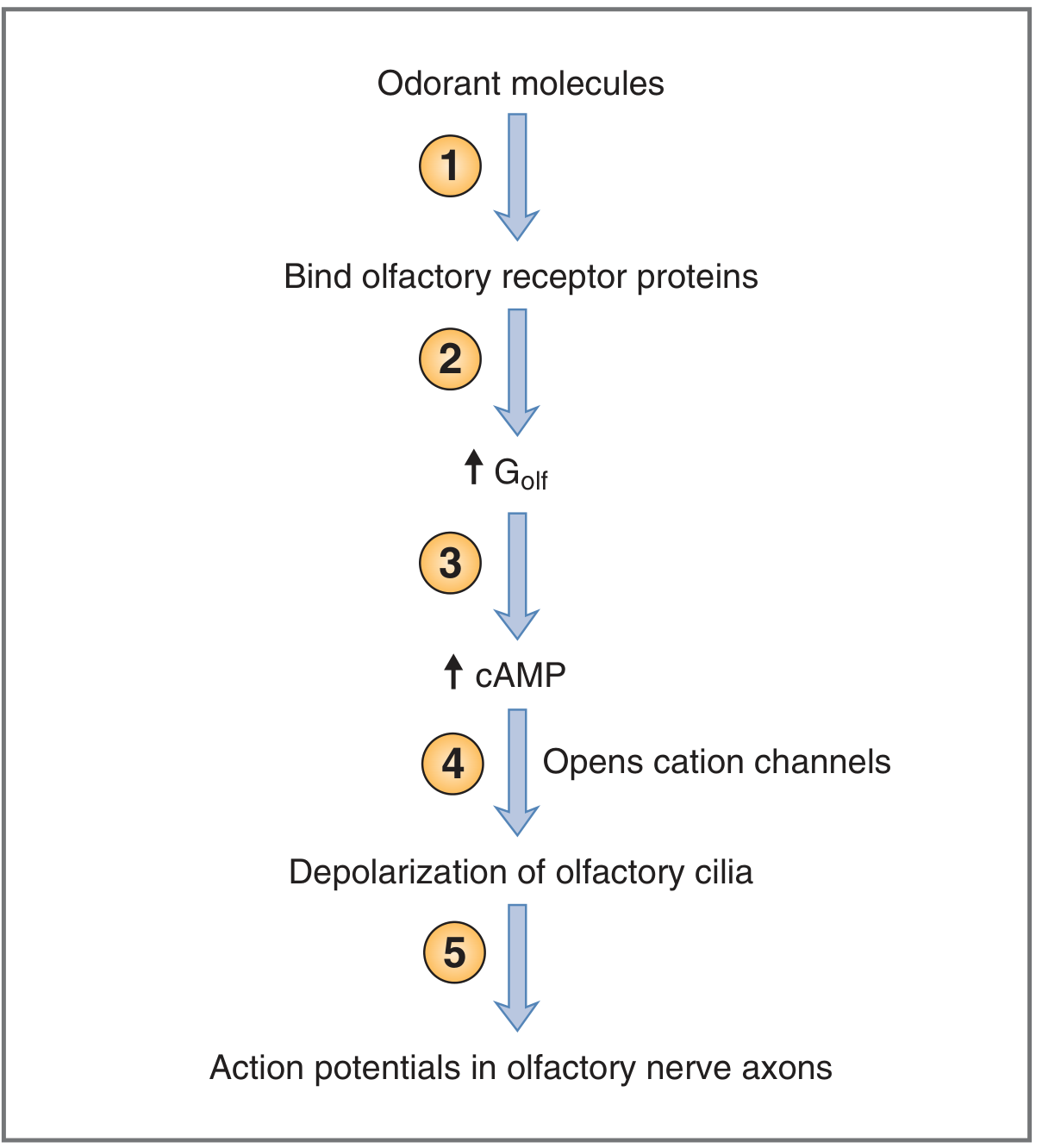
Fig. 4 - Steps in olfactory transduction. (Costanzo Physiology 7e, Fig. 3.27)
Step-by-step mechanism (Costanzo; Guyton & Hall; Ganong; Boron & Boulpaep):
ODORANT (volatile, water-soluble, lipid-soluble molecule)
|
▼
Diffuses through mucous layer
(assisted by Odorant-Binding Proteins - OBPs)
|
▼
Binds to specific OLFACTORY RECEPTOR PROTEIN on cilia
(7-transmembrane domain GPCR; ~400 functional types in humans)
|
▼
Conformational change → activation of G-protein (G_olf / G_αII)
|
▼
G_olf α-subunit dissociates → activates ADENYLYL CYCLASE (AC-III)
|
▼
AC-III converts ATP → ↑↑↑ cAMP (second messenger; signal amplification)
|
▼
cAMP binds to CYCLIC NUCLEOTIDE-GATED (CNG) CATION CHANNELS
(permeable to Na⁺, K⁺, Ca²⁺)
|
▼
CNG channels OPEN → Net inward current → MEMBRANE DEPOLARIZATION
(raised from resting -55 mV toward -30 mV)
|
▼
↑ Intracellular [Ca²⁺] → opens Ca²⁺-activated Cl⁻ channels (ANOCTAMIN-2/ANO2)
→ further depolarization (due to high intracellular [Cl⁻] in ORN)
|
▼
If threshold exceeded → ACTION POTENTIALS generated in ORN axon
|
▼
Propagated along unmyelinated CN I fibers → OLFACTORY BULB
Diagram 4: Molecular mechanism of olfactory transduction (Guyton & Hall, Fig. 54.4)
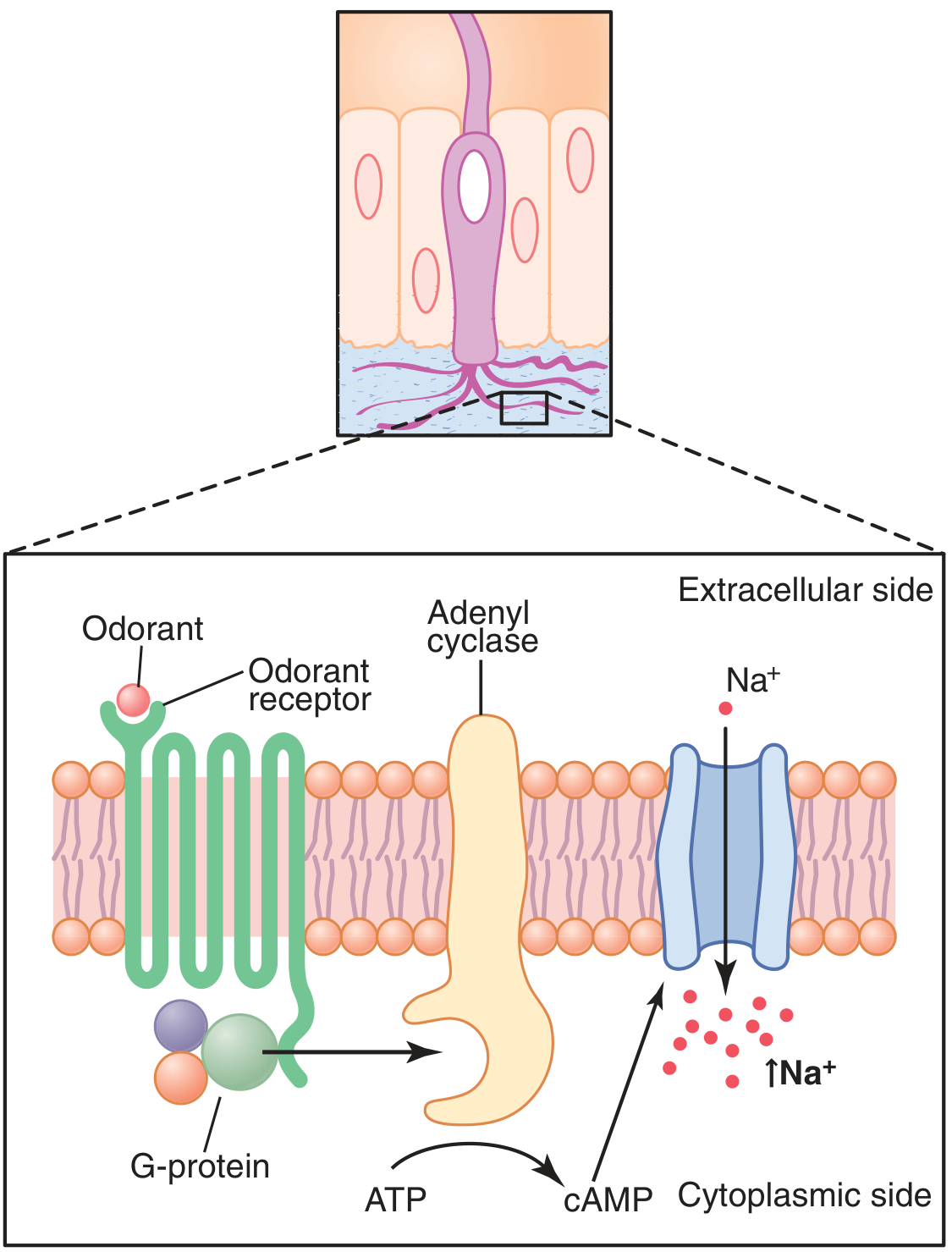
Fig. 5 - Summary of olfactory signal transduction. (Guyton & Hall, Fig. 54.4)
Diagram 5: G-protein coupled olfactory transduction (Ganong's Review, Fig. 9-2)
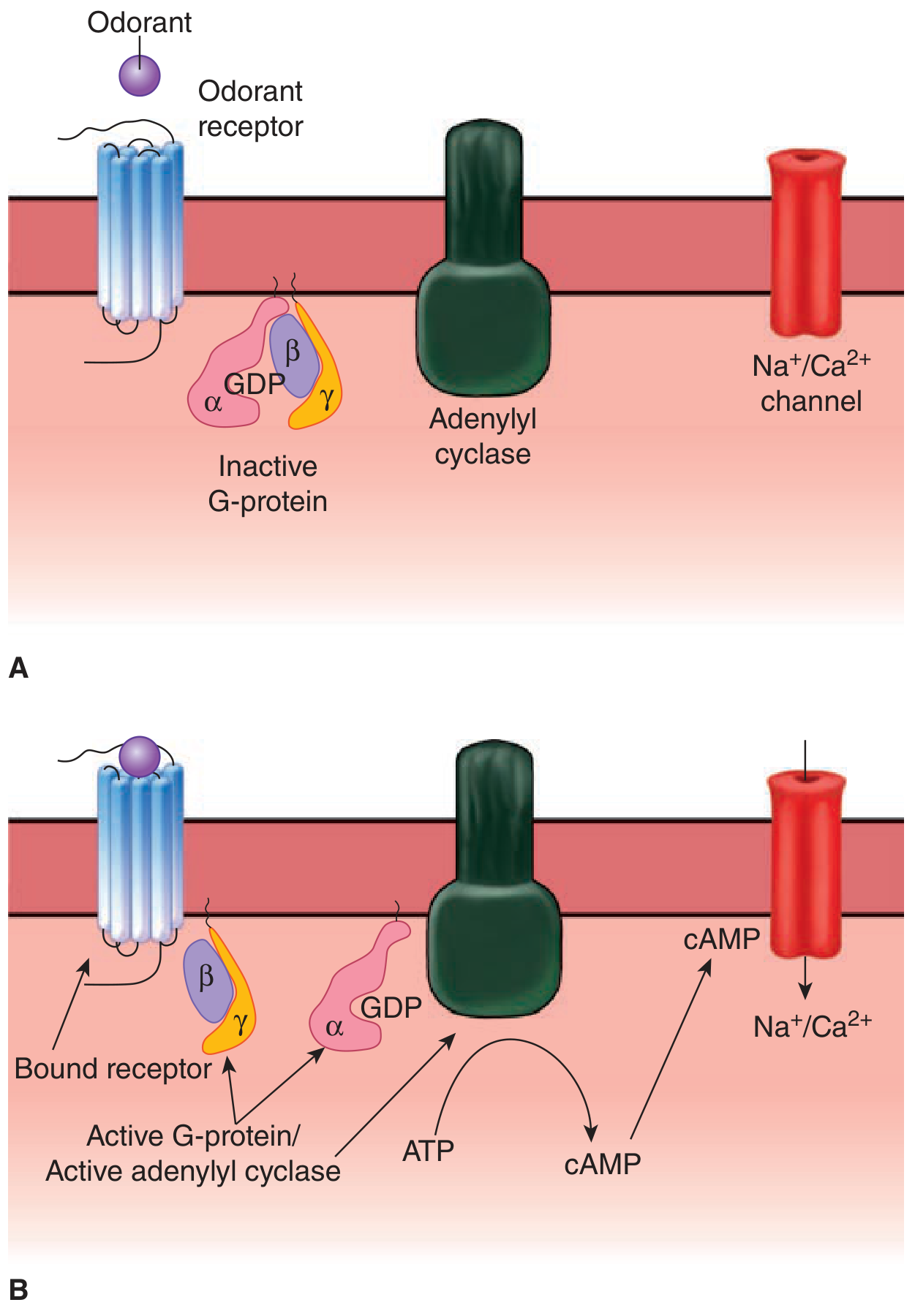
Fig. 6 - G-protein coupled odorant receptor signalling showing A) inactive state with G-protein bound to GDP, and B) active state where alpha subunit activates adenylyl cyclase to generate cAMP. (Ganong's Review, Fig. 9-2)
Signal Amplification (Guyton & Hall)
This cascade is a three-step amplification system:
- One receptor protein activates multiple G-protein molecules
- Each G-protein activates multiple adenylyl cyclase molecules
- Each adenylyl cyclase produces many cAMP molecules, opening many CNG channels
Result: Even a single odorant molecule can trigger detectable neural activity. The sensitivity of olfaction to trace concentrations is explained entirely by this cascade amplification.
Adaptation (Guyton & Hall)
- Olfactory receptors adapt ~50% within the first second
- Further adaptation (near-extinction of perception) occurs at the central nervous system level via centrifugal inhibitory fibers from the brain back to the OB
Prerequisites for Olfactory Stimulation (Guyton & Hall)
- Substance must be volatile (in gas phase)
- Must be at least slightly water-soluble (to diffuse through mucus)
- Must be at least slightly lipid-soluble (to penetrate ciliary membrane)
PART III: CLASSICAL THEORIES OF OLFACTION
Before the molecular mechanism was established, several competing theories were proposed to explain odor perception.
THEORIES OF OLFACTION
├── Physical Theories
│ ├── Vibration/Infrared Theory (Dyson 1928, Wright 1954, Turin 1996)
│ └── Puncturing Theory (Müller)
├── Chemical/Molecular Shape Theories
│ ├── Stereochemical Theory (Moncrieff 1949, Amoore 1952)
│ ├── Profiling Theory (Beets, 1970s)
│ └── Enzyme Induction Theory
├── Modern Receptor Theories
│ ├── Lock-and-Key / GPCR Theory (Buck & Axel 1991) ← Currently accepted
│ ├── Combinatorial Receptor Code Theory
│ └── Spatial Map Theory (Glomerular map)
└── Neural Coding Theories
├── Across-Fiber Pattern Theory
└── Temporal Coding Theory
THEORY 1: VIBRATION THEORY (Dyson 1937; Wright 1954; Turin 1996)
Proposed by: G. Malcolm Dyson (1928, 1937), expanded by R.H. Wright (1954), revived by Luca Turin (1996).
Core Concept: The smell character of a molecule is determined by its intramolecular vibrational frequency in the infrared range, rather than (or in addition to) its shape.
Historical progression:
- Ogle (1870): Suggested an "undulatory theory" of smell analogous to wave theories of light and sound
- Dyson (1928-1938): Proposed that vibrational modes within the range 1,400-3,500 cm⁻¹ activate olfactory receptor sites. Called the "Raman effect" theory
- Wright (1954): Extended the concept using quantum and thermodynamic considerations; correlated Raman spectra with odor character
- Turin (1996): Revived the theory, proposing that G-protein-coupled olfactory receptors detect molecular vibrations through inelastic electron tunneling - the "swipe card" model
Turin's Mechanism ("Swipe Card" Model):
Odorant molecule enters receptor binding site (shape-dependent)
↓
IF vibrational energy mode matches energy gap between
two receptor energy levels
↓
Electron tunnels THROUGH the molecule (inelastic electron tunneling)
↓
Energy transfer → triggers G-protein signaling cascade
↓
Odor perception
Evidence for: Deuterated molecules (same shape, different vibrational frequency) sometimes smell different; fruit flies can distinguish deuterated from non-deuterated odorants.
Evidence against: Keller & Vosshall (2004) found humans could NOT reliably distinguish deuterated from normal acetophenone. Turin's proposed tunneling mechanism challenged on biophysical grounds. The theory remains controversial and unproven.
Current status: The vibration theory is a minority view. The shape/receptor theory is dominant. Stoneham et al. (UCL, 2007) showed Turin's mechanism was at least physically plausible ("consistent with known physics"), but biological evidence is lacking.
THEORY 2: STEREOCHEMICAL THEORY (Amoore 1952; Moncrieff 1949)
Proposed by: R.W. Moncrieff (1949), J.E. Amoore (1952, 1963-1964). This is the Lock-and-Key theory of olfaction.
Core Concept: The odor quality of a substance is determined by the 3-dimensional geometric shape and size of its molecule, which must fit into a complementary receptor site (like a key fitting a lock).
Amoore's Primary Odors (1952):
Amoore analyzed 600 organic compounds described in chemical literature and identified 7 primary odors, each with a specific molecular shape:
| Primary Odor | Molecular Shape | Example |
|---|---|---|
| Camphoraceous | Spherical | Camphor, cyclohexane |
| Musky | Disc-shaped | Pentadecanolide |
| Floral | Disc with tail | Geraniol, phenylethanol |
| Peppermint | Wedge-shaped | Menthol |
| Ethereal | Rod-shaped, small | Diethyl ether |
| Pungent | Electrophilic | Formic acid |
| Putrid | Nucleophilic | Hydrogen sulfide |
Evidence for:
- Enantiomers (mirror-image molecules, identical chemistry but opposite geometry) often smell different - e.g., R-(-)-carvone smells of spearmint, S-(+)-carvone smells of caraway
- Specific anosmia: individuals may lack sensitivity to specific odors while smelling others normally - suggests dedicated receptor sites
Limitations:
- Cannot explain the vast diversity of human olfactory discrimination (>1 million odors, not just 7 primaries)
- Fails to account for molecules with similar shapes but different smells
- The concept of only 7 primaries has been abandoned; modern understanding recognizes ~400 receptor types
Legacy: The stereochemical theory was the dominant framework until the molecular biology revolution of the 1990s. The fundamental concept - that molecular geometry determines receptor binding - is validated in the modern GPCR receptor theory.
THEORY 3: PROFILING / FUNCTIONAL GROUP THEORY (Beets, 1970s)
Proposed by: M.G.J. Beets.
Core Concept: Odor character is determined not just by molecular shape, but by the combination of:
- The profile (overall molecular shape/framework)
- The functional group (the chemically reactive portion of the molecule)
Beets argued that neither the shape alone nor the functional group alone determines the smell - both together create the odor "profile."
Example: Musks all share a similar macrocyclic or polycyclic framework but differ in functional groups, which modulates the exact character of the musk odor.
Current relevance: This theory is supported by modern receptor-binding data - the binding pocket of olfactory receptors interacts with both the backbone structure AND specific chemical moieties of odorant molecules.
THEORY 4: ENZYME INDUCTION THEORY (Davies & Taylor)
Proposed by: J.T. Davies and F.H. Taylor (1959).
Core Concept: Odorant molecules, by virtue of their ability to penetrate the lipid receptor membrane based on their partition coefficient (oil/water partition), pierce the receptor membrane and induce a conformational change in membrane proteins (enzymes). This enzyme activation generates an electrical signal.
Mechanism proposed:
Odorant in mucous layer
↓
Penetrates lipid membrane of cilia (proportional to lipid solubility)
↓
Punctures/disrupts membrane → induces enzyme conformational change
↓
Enzymatic activity generates ionic current
↓
Depolarization → action potentials
Supporting observation: There is a rough correlation between lipid solubility of odorants and their detection threshold - more lipid-soluble compounds generally have lower thresholds.
Limitations: Cannot explain odor specificity or discrimination; all lipid-soluble molecules should smell the same under this theory - which they do not.
THEORY 5: MODERN RECEPTOR THEORY - COMBINATORIAL CODE (Buck & Axel 1991) - CURRENTLY ACCEPTED
Proposed by: Linda Buck and Richard Axel (1991) - awarded the Nobel Prize in Physiology or Medicine, 2004.
Core Concept: There are ~400 functional olfactory receptor genes in humans (>1100 in rodents; ~2000 in elephants), each encoding a distinct 7-transmembrane domain GPCR. Odorant recognition is based on a combinatorial code:
COMBINATORIAL RECEPTOR CODE:
- Each ORN expresses ONLY ONE type of receptor gene
- Each receptor responds to MULTIPLE odorants (with varying affinity)
- Each odorant activates MULTIPLE receptors (with varying affinities)
- The PATTERN of receptor activation = the odor "code"
Evidence from Neuroscience: Exploring the Brain (5th Ed.):
- ~400 different receptor genes make this by far the largest family of mammalian genes (3-5% of entire genome)
- Mice: >1,100 receptor types; African elephants: ~2,000
- Nearly all odorants activate more than one receptor
- Nearly all receptors respond to multiple odorants
- Each glomerulus in the OB receives input from ORN expressing the same receptor type - creating a spatial/combinatorial map
Diagram 6: Flow chart of Combinatorial Receptor Code Theory:
~400 Olfactory Receptor (OR) Genes
↓
Each ORN expresses ONE OR gene → ONE OR type
↓
Odorant molecule binds to multiple ORs with different affinities
↓
Pattern of activated ORs = COMBINATORIAL CODE for that odorant
↓
ORNs with same OR type converge on same GLOMERULUS in OB
↓
Spatial pattern of activated glomeruli = GLOMERULAR MAP
↓
Mitral/tufted cells relay unique spatial-combinatorial pattern
↓
Higher olfactory cortex (piriform cortex) decodes the pattern
↓
ODOR PERCEPTION / IDENTIFICATION
This theory explains:
- How humans can distinguish >1 million odors with only 400 receptor types (combinatorial mathematics: 2^400 = astronomically large number of possible patterns)
- Specific anosmia: mutation or absence of one OR gene → loss of sensitivity to those specific odorants
- Odor quality perception
THEORY 6: SPATIAL MAP THEORY (Glomerular Map)
Basis: Each OR type is mapped to a specific glomerulus in the OB. ORNs expressing the same OR gene (regardless of where in the OE they are located) all project to the same 1-2 glomeruli in the OB. This creates a reproducible topographic odor map in the OB.
Key finding (Neuroscience: Exploring the Brain):
- One odor activates a particular combination of glomeruli
- One glomerulus (or mitral cell) is activated by multiple odorants
- The "map" is preserved across individuals of the same species
Criticism (Neuroscience textbook): Spatial maps may be a developmental byproduct of efficient wiring (minimizing axon/dendrite length) rather than the primary mechanism for encoding odor identity. The olfactory cortex has not been shown to "read" spatial maps the way vision reads retinotopic maps.
THEORY 7: TEMPORAL CODING THEORY
Basis: Growing evidence that the temporal patterns of spiking in olfactory neurons encode odor quality, not just spatial maps.
Evidence (Neuroscience: Exploring the Brain, Fig. 8.23):
- The olfactory bulb and cortex generate oscillations when odors are presented
- Odors change not just which cells fire but when they fire within the respiratory cycle
- Odor 1 may change spike timing without changing spike number
- Odor 2 may increase spike number without changing timing
- Odor 3 may both increase spike number and delay onset
Dhawale et al. (2010, Nature Neuroscience): Demonstrated non-redundant odor coding by sister mitral cells using light-addressable glomeruli - different odors recruit different temporal patterns even from the same cell.
THEORY 8: ACROSS-FIBER PATTERN THEORY
Concept: No single receptor or fiber is uniquely dedicated to a single odor. Rather, olfactory coding is distributed across many fibers simultaneously. The pattern of activity across the entire ensemble of olfactory neurons encodes the odor - analogous to how color vision uses overlapping spectral sensitivities of three cone types.
This theory reconciles with the combinatorial code and is now largely integrated into modern understanding.
PART IV: OLFACTORY CODING - HOW ARE ODORS ENCODED?
Summary Table: Theories Compared
| Theory | Proponent | Year | Basis | Status |
|---|---|---|---|---|
| Vibration/IR | Dyson, Wright, Turin | 1928-1996 | Molecular vibrational frequency | Controversial, minority view |
| Stereochemical/Lock-Key | Moncrieff, Amoore | 1949-1963 | Molecular 3D shape | Partially validated; superseded |
| Profiling | Beets | 1970s | Shape + functional group | Partially valid |
| Enzyme/Puncturing | Davies & Taylor | 1959 | Membrane penetration | Largely abandoned |
| GPCR Combinatorial Code | Buck & Axel | 1991 | 400 receptor gene families | Currently accepted |
| Spatial/Glomerular Map | Multiple | 1990s-2000s | Topographic OB map | Valid but may be developmental artifact |
| Temporal Coding | Multiple | 2000s | Spike timing patterns | Emerging evidence |
| Across-Fiber Pattern | Multiple | 1970s+ | Population coding | Valid; integrated with above |
PART V: CLASSIFICATION OF OLFACTORY DYSFUNCTION
(Cummings Otolaryngology, Table 36.1)
| Term | Definition |
|---|---|
| Normosmia | Normal olfactory function |
| Hyposmia (microsmia) | Quantitatively reduced olfaction |
| Functional anosmia | Reduced to the extent of no useful function in daily life |
| Anosmia | Complete absence of olfactory function |
| Specific anosmia | Loss for one specific odorant only (supports receptor theory) |
| Hyperosmia | Abnormally heightened smell (rare; associated with migraine) |
| Parosmia (dysosmia) | Distorted perception in presence of odorant |
| Phantosmia | Olfactory hallucination without stimulus |
| Orthonasal olfaction | Perception via anterior airflow (sniffing) |
| Retronasal olfaction | Perception via posterior airflow during swallowing (basis of flavor) |
PART VI: RECENT ADVANCES
1. Nobel Prize - Buck & Axel (2004)
The discovery of olfactory receptor genes (1991) - the largest mammalian gene family - transformed our understanding of olfactory coding. Over 400 functional OR genes in humans, each expressed in a unique ORN type, each projecting to a unique glomerulus. [Manzini, Schild, Di Natale - Physiological Reviews 2022; 102(1):61-154]
2. Odorant-Binding Proteins (OBPs)
Hydrophilic carrier proteins in nasal mucus that facilitate transport of hydrophobic odorants to ORs. Also serve to terminate the olfactory signal by removing odorants from receptors. Role in olfactory adaptation.
3. Olfactory Ensheathing Cells (OECs)
Unique glial cells in the OE and OB that guide ORN axon regeneration. Autologous OEC transplantation has been investigated as a therapeutic strategy for nerve injury (Cummings). Stem cell and regeneration research is ongoing.
4. COVID-19 and Olfactory Dysfunction
The SARS-CoV-2 pandemic produced a surge of research into olfactory mechanisms. Butowt et al. (Trends in Neuroscience, 2023; PMID: 36470705) demonstrated that anosmia in COVID-19 results from virus infecting sustentacular (supporting) cells rather than ORN directly, disrupting the microenvironment and causing ORN dysfunction - consistent with the known biology of the supporting cells in the OE.
5. Olfactory Training
Repeated, structured exposure to defined odorants (rose, eucalyptus, lemon, clove) - "olfactory training" - has shown efficacy in post-viral and post-traumatic olfactory loss by stimulating neuroregeneration. Supported by evidence in Cummings (2020).
6. Non-olfactory Odorant Receptors
OR genes are expressed in many non-olfactory tissues (gut, heart, testes, kidney). Gut odorant receptors modulate hormone secretion (Wang et al., Crit Rev Food Sci Nutr, 2024; PMID: 36785901) - suggesting olfactory receptors have broader physiological roles.
7. Trace Amine-Associated Receptors (TAARs)
A newly discovered small family of olfactory receptors (not GPCRs of the classic OR gene family). TAARs respond to substances released by protein decomposition and food spoilage. Expressed in olfactory receptor cells but play a relatively minor role in human olfaction.
8. AI and Machine Olfaction
Recent machine learning models (Barwich & Lloyd, Front. Neurosci. 2022; PMID: 36117640) attempt to predict odor character from molecular structure. These models have validated the combinatorial receptor code theory computationally but also revealed its limits - structural prediction of odor quality remains imperfect, suggesting undiscovered variables in the olfactory system.
9. Receptor Structure Decoding (2026)
Lu, Vihani, Nagai et al. (Research Square, 2026; PMID: 42326533 - preprint) - "Decoding Smell from Receptor Structure" represents the frontier of direct receptor-odorant binding structure analysis using cryo-EM, potentially answering questions about the structural basis of odor quality prediction at atomic resolution.
SUMMARY FLOW CHART: COMPLETE OLFACTORY PATHWAY
ODORANT (volatile, water-soluble, lipid-soluble molecule in air)
│
▼
Enters nasal cavity → turbinates create turbulent flow
│
▼
Reaches olfactory cleft (upper nasal cavity)
│
▼
Dissolves in MUCOUS LAYER (assisted by Odorant-Binding Proteins)
│
▼
Binds OLFACTORY RECEPTOR (7-TM GPCR on cilia of ORN)
│
▼
G_olf activation → Adenylyl Cyclase → ↑ cAMP → CNG channels open
│
▼
Membrane depolarization → ACTION POTENTIALS in CN I fibers
│
▼
Unmyelinated axons pass through CRIBRIFORM PLATE foramina
│
▼
Synapse in GLOMERULI of OLFACTORY BULB
(same OR type → same glomerulus → spatial map)
│
▼
MITRAL/TUFTED CELLS (2nd order neurons)
[modulated by periglomerular and granule cell lateral inhibition]
│
▼
LATERAL OLFACTORY STRIA
│
▼
PRIMARY OLFACTORY CORTEX
(Piriform cortex, anterior olfactory nucleus, olfactory tubercle,
amygdala, entorhinal cortex)
│
▼
ORBITOFRONTAL CORTEX (via thalamus)
= Conscious odor perception + quality discrimination
│
(amygdala connection)
▼
EMOTIONAL / MEMORY ASSOCIATIONS (limbic system)
REFERENCES (from textbooks consulted)
- Scott-Brown's Otorhinolaryngology Head & Neck Surgery, Vol. 1, Ch. on Nasal Physiology & Olfaction
- Cummings Otolaryngology Head & Neck Surgery, 7th Ed., Ch. 36 (Olfactory Neuroepithelium, Olfactory Dysfunction)
- Costanzo Physiology, 7th Ed., pp. 104-107 (Olfaction, Transduction, Encoding)
- Guyton & Hall Textbook of Medical Physiology, pp. 671-673 (Sense of Smell)
- Ganong's Review of Medical Physiology, 26th Ed., pp. 187-189 (Odorant Receptors, Signal Transduction, Olfactory Pathway)
- Boron & Boulpaep Medical Physiology, pp. 533-534 (Olfactory Transduction - detailed molecular steps including ANO2)
- Neuroscience: Exploring the Brain, 5th Ed., pp. 787-810 (Receptor proteins, Combinatorial code, Spatial maps, Temporal coding)
- Manzini I, Schild D, Di Natale C. Principles of odor coding in vertebrates and artificial chemosensory systems. Physiol Rev. 2022;102(1):61-154
- Butowt R, Bilinska K, von Bartheld CS. Olfactory dysfunction in COVID-19. Trends Neurosci. 2023;46(1). [PMID: 36470705]
- Amoore JE. Stereochemical theory of olfaction. Nature. 1963;198:271-272
- Turin L. A spectroscopic mechanism for primary olfactory reception. Chem Senses. 1996;21:773-791
- Hoehn RD et al. Status of the Vibrational Theory of Olfaction. Front Physics. 2018;6:25
RGUHS Exam Tip: For a 20-mark answer, RGUHS expects: (1) Introduction with importance of olfaction, (2) Anatomy of olfactory epithelium and pathway with labeled diagrams, (3) All classical theories (stereochemical, vibration, enzyme induction) with proponents and evidence, (4) Modern GPCR/combinatorial code theory as the accepted mechanism with detailed transduction steps and flow charts, (5) Recent advances including Nobel Prize (Buck & Axel), COVID-19 anosmia, olfactory training, and OECs. Dhingra's and Hazarika's ENT textbooks present the same classical theories (stereochemical, vibration, enzyme/chemical) as described above, and the accepted modern GPCR mechanism - the content above covers all these comprehensively. Zakir Hussain's ENT notes follow the same framework.
This is a shared conversation. Sign in to Orris to start your own chat.