Cerebellum
Reading File
Asking for Preferences
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Reading File
Loading Image
Reading File
I now have comprehensive content from multiple authoritative textbooks. Let me compile the full answer.
Cerebellum: Physiology & Circuits
Overview
The cerebellum sits in the posterior cranial fossa, dorsal to the pons and medulla. Despite comprising only ~10% of brain volume, it contains more than half of all neurons in the brain. Its function is to coordinate movement, plan and execute motor sequences, maintain posture, and coordinate head and eye movements - acting as an integrator between the cerebral cortex, spinal cord, and vestibular organs.
"The cerebellum, conveniently positioned between the cerebral cortex and the spinal cord, integrates sensory information about position from the spinal cord, motor information from the cerebral cortex, and information about balance from the vestibular organs of the inner ear."
- Costanzo Physiology 7th Ed.
Functional Longitudinal Zones
The cerebellum is divided into three functional zones based on longitudinal position, each with distinct inputs, outputs, and functions:
| Zone | Anatomical Region | Deep Nucleus | Function |
|---|---|---|---|
| Vestibulocerebellum | Flocculonodular lobe | Vestibular nuclei (direct) | Balance, vestibulo-ocular reflex, smooth-pursuit eye movements |
| Spinocerebellum | Vermis + intermediate hemisphere | Fastigial / Interposed nuclei | Axial & limb muscle coordination; correction of ongoing movement |
| Cerebrocerebellum (Pontocerebellum) | Lateral hemisphere | Dentate nucleus | Planning & initiation of voluntary movement; cognitive functions |
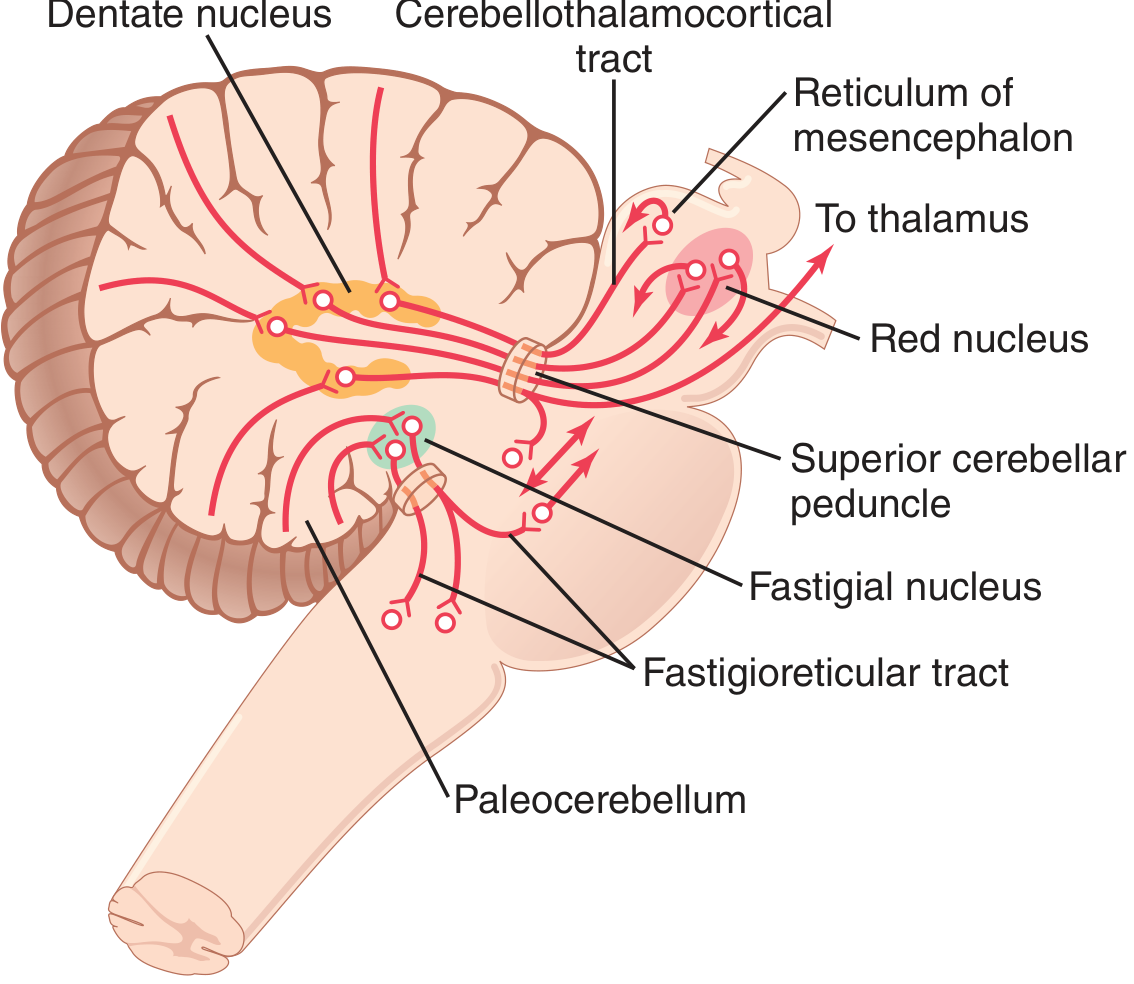
The vermis projects via the fastigial nucleus to brain stem and spinal cord (via the fastigioreticular tract) controlling axial/proximal muscles. The intermediate hemisphere projects via the interposed nucleus (globose + emboliform) to the red nucleus and motor cortex to control distal limb muscles. The lateral hemisphere projects via the dentate nucleus to the ventrolateral thalamus and then the motor and premotor cortex for planning sequential voluntary movements.
Cerebellar Cortex - Three Layers
The cortex has three layers and five cell types:
- Molecular layer (outer) - contains stellate cells, basket cells, and the dendrites of Purkinje cells and axons of granule cells (parallel fibers)
- Purkinje cell layer (middle) - a single cell-thick layer of the largest neurons in the CNS
- Granular layer (inner) - densely packed granule cells and Golgi cells
The Canonical Microcircuit
This repeating unit (about 30 million copies) is the fundamental computational element of the cerebellum:
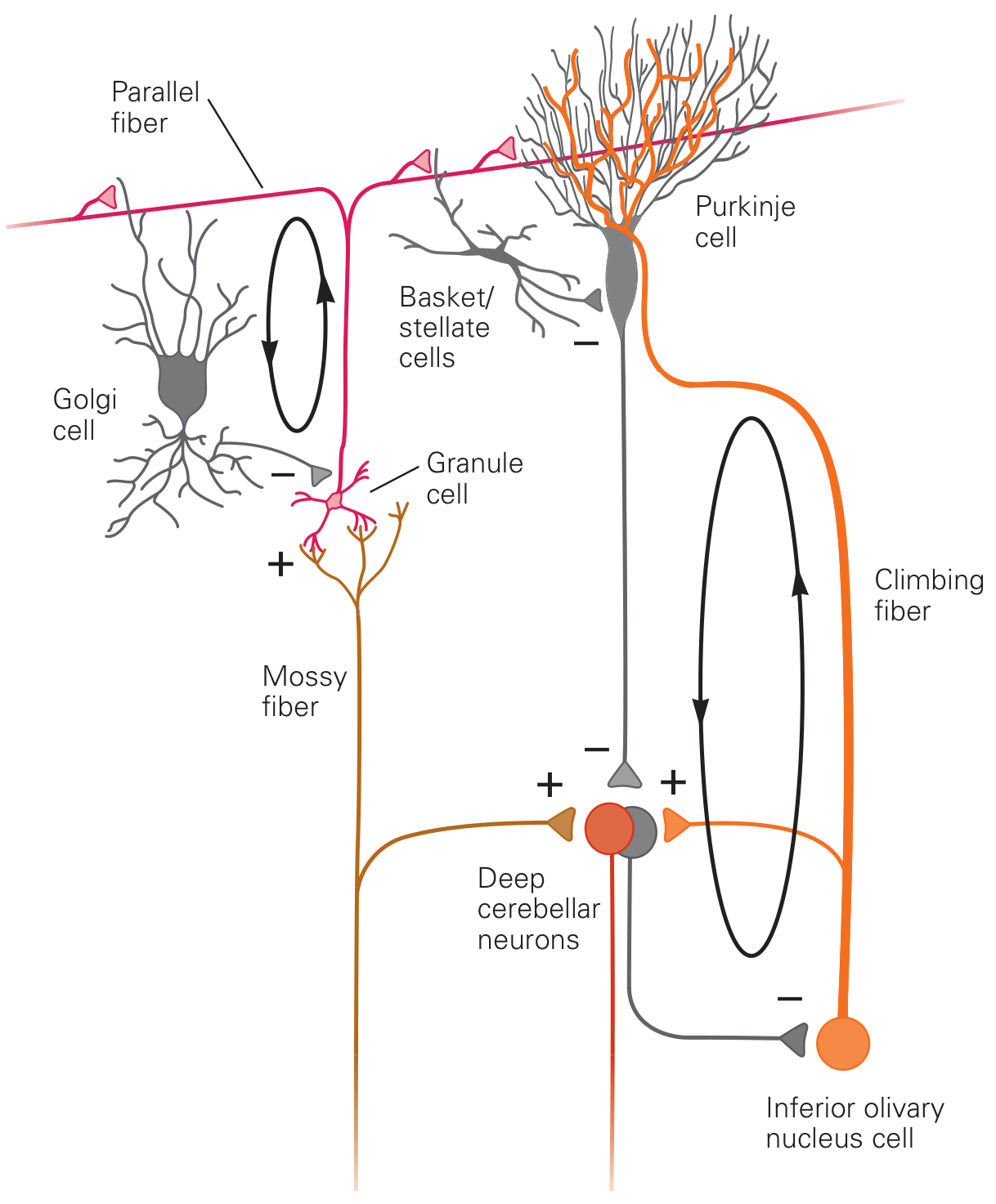
Two Afferent Fiber Systems
1. Mossy Fibers
- Originate from the spinal cord, pontine nuclei, vestibular nuclei, and brainstem reticular formation
- Enter via the inferior and middle cerebellar peduncles
- Send collaterals to deep cerebellar nuclei (excitatory, +)
- Synapse on granule cells in the granular layer
- Granule cell axons ascend and bifurcate into parallel fibers in the molecular layer - each running 1-2 mm in each direction
- Each Purkinje cell receives input from 80,000-200,000 parallel fibers
- Produce simple spikes in Purkinje cells (weak, graded, high-frequency modulation)
2. Climbing Fibers
- Originate exclusively from the inferior olivary nucleus of the medulla
- One climbing fiber wraps around 5-10 Purkinje cells and makes ~300 synapses per cell
- Also send collaterals to deep cerebellar nuclei (excitatory, +)
- Produce complex spikes in Purkinje cells - powerful, all-or-nothing, low frequency (~1 Hz)
- Function as an error signal for motor learning
Inhibitory Interneurons in the Cortex
- Basket cells: axons run perpendicular to parallel fibers; make powerful inhibitory synapses on Purkinje cell bodies → lateral inhibition (sharpening of signals)
- Stellate cells: act locally on Purkinje cell dendrites
- Golgi cells: in the granular layer; receive excitatory input from parallel fibers and mossy fibers → feed back inhibition onto granule cells (recurrent loop)
Output - The Purkinje Cell
- Purkinje cells are the sole output of the cerebellar cortex
- They project to the deep cerebellar nuclei (and directly to vestibular nuclei)
- Purkinje cells are GABAergic (inhibitory)
- They fire tonically at 50-100 action potentials/sec at rest
- Deep cerebellar nuclear cells are simultaneously excited by mossy/climbing fiber collaterals and inhibited by Purkinje cells
- The net output from deep nuclei is determined by the balance of these two influences
Turn-On/Turn-Off Mechanism
This is the signature physiological output of the cerebellum (Guyton & Hall):
When a rapid movement is initiated, deep nuclear cells first receive excitatory drive from mossy/climbing fiber collaterals → a burst of output. Milliseconds later, Purkinje cell inhibition arrives (via the granule → parallel fiber → Purkinje cell delay pathway) → a damping inhibitory signal. This prevents overshoot and oscillation of the movement.
This creates:
- Turn-ON signal for agonist muscles at movement onset
- Turn-OFF signal for agonist + reciprocal turn-on for antagonist at movement termination
Computational Functions (Kandel)
The cerebellum is hypothesized to perform four key computational operations:
1. Feedforward Sensorimotor Control
Sensory feedback is inherently delayed. The cerebellum pre-programs muscle contractions before sensory feedback arrives, anticipating what forces will be needed. Cerebellar neurons fire before movement begins, unlike spinal reflex arcs.
2. Internal Models
The cerebellum builds and maintains two types of internal models:
- Inverse dynamic model: processes current limb posture → generates properly timed muscle commands
- Forward dynamic model: takes a copy of the motor command (efference copy) → predicts upcoming movement kinematics
These allow the cerebellum to compensate for interaction torques between linked body segments (e.g., elbow flexion causing unintended shoulder movement). Cerebellar patients fail at this - a hallmark of ataxia.
3. Integration of Sensory Input with Corollary Discharge
The cerebellum compares corollary discharge (internal copy of motor commands from the motor cortex) with incoming sensory signals about actual movement, to detect and correct errors in real time.
4. Timing Control
The cerebellum is involved in sub-second timing of motor sequences. The repeating microcircuit architecture, with its precise delays through parallel fiber systems, is thought to provide the temporal framework for coordinating muscle contractions.
Cerebellar Peduncles - I/O Routes
| Peduncle | Direction | Contents |
|---|---|---|
| Superior (brachium conjunctivum) | Mainly efferent | Dentate/interposed → red nucleus, thalamus |
| Middle (brachium pontis) | Afferent only | Pontocerebellar fibers (contralateral pontine nuclei) |
| Inferior (restiform body) | Mixed | Afferent: spinocerebellar, olivocerebellar, vestibulocerebellar; Efferent: to vestibular nuclei |
Motor Learning
The Marr-Albus hypothesis (1970s): the climbing fiber from the inferior olive acts as a teaching signal, encoding movement errors. When a climbing fiber fires coincidently with parallel fiber activation of a Purkinje cell, it causes long-term depression (LTD) of that parallel fiber synapse. Over repeated trials, this selectively weakens the synaptic connections associated with erroneous movements, thereby improving performance. This is the cellular substrate for cerebellar motor learning.
Key evidence:
- Climbing fibers fire at low rates but with high precision, triggered by sensory prediction errors
- Flocculus (vestibulocerebellum) damage abolishes VOR adaptation
- Inactivation of the interposed nucleus blocks conditioned eyeblink learning
Summary of Key Physiology Points
| Feature | Detail |
|---|---|
| Sole cortical output | Purkinje cell (inhibitory, GABAergic) |
| Climbing fiber origin | Inferior olivary nucleus only |
| Mossy fiber origins | Spinal cord, pontine nuclei, vestibular nuclei, reticular formation |
| Purkinje cell resting rate | 50-100 Hz |
| Granule:Purkinje ratio | ~500-1000:1 |
| Parallel fiber synapses per Purkinje cell | 80,000-200,000 |
| Deep nuclei (medial → lateral) | Fastigial → Interposed (globose + emboliform) → Dentate |
| Climbing fiber spike type | Complex spike (low freq, ~1 Hz) |
| Mossy fiber spike type | Simple spike (high freq, graded) |
Sources: Guyton & Hall Textbook of Medical Physiology; Ganong's Review of Medical Physiology 26th Ed.; Kandel, Principles of Neural Science 6th Ed.; Costanzo Physiology 7th Ed.
This is a shared conversation. Sign in to Orris to start your own chat.