Explain the biochemistry of pancreatic secretions
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Reading File
Loading Image
I now have excellent content from multiple authoritative textbooks and detailed diagrams. Let me write the comprehensive answer.
Biochemistry of Pancreatic Secretions
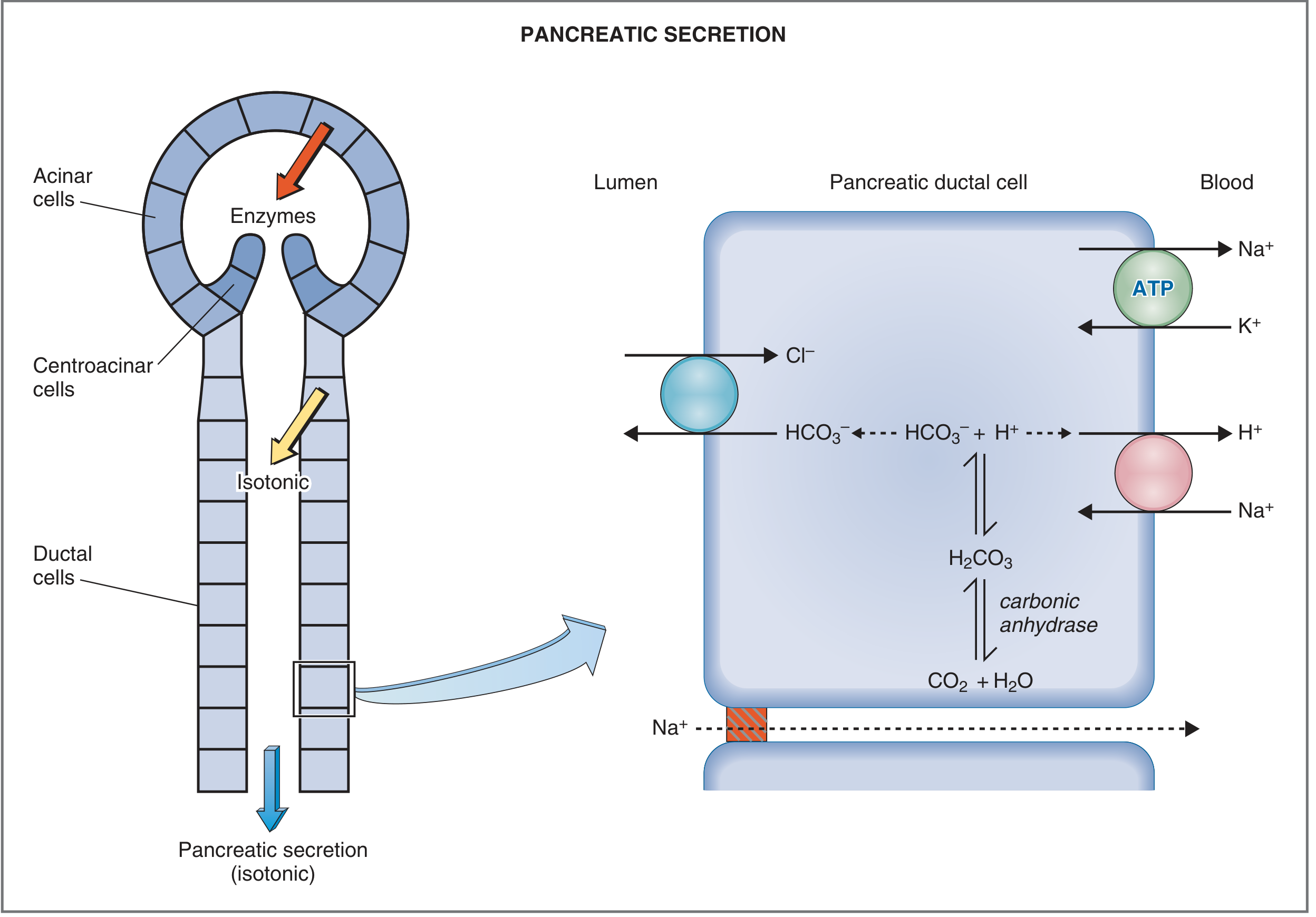
The exocrine pancreas produces approximately 1-2.5 L of fluid per day - an isotonic, alkaline juice containing two distinct components: an aqueous (bicarbonate-rich) component from ductal/centroacinar cells, and an enzymatic component from acinar cells. These are produced by entirely separate biochemical mechanisms.
1. Functional Anatomy Overview
The exocrine pancreas is organized like a bunch of grapes. Each acinus (from Latin "berry in a cluster") is the secretory blind end of a branching duct system, lined by acinar cells. Ductal epithelial cells extend into the acinus as centroacinar cells. The ductule drains into interlobular ducts, then into the main pancreatic duct.
- Acinar cells: basal nucleus, abundant rough ER, apical zymogen granules - specialized for protein synthesis and enzyme secretion
- Centroacinar/ductal cells: abundant mitochondria, contain carbonic anhydrase - specialized for ion transport and bicarbonate secretion
2. The Enzymatic Component (Acinar Cells)
2a. Synthesis and Intracellular Transport
Pancreatic digestive enzymes follow the classical secretory protein pathway:
- Synthesis on rough ER - mRNAs for digestive enzymes are translated on membrane-bound ribosomes and co-translationally inserted into the ER lumen
- Transfer to Golgi complex - proteins are processed and glycosylated; sorting signals direct them to secretory vs. lysosomal pathways. Lysosomal enzymes receive mannose-6-phosphate tags in the cis-Golgi; digestive enzymes are directed by the pancreas consensus element (a 5' enhancer region) regulated by pancreas transcription factor-1 (PTF1), which is selectively expressed in the exocrine pancreas
- Condensing vacuoles - enzymes concentrate into zymogen granules at the apical region of the cell
- Exocytosis - stimulated granules migrate to the apical plasma membrane via an actin-myosin cytoskeletal network; SNARE proteins and GTP-binding proteins mediate membrane fusion and release into the acinar lumen
2b. Enzyme Inventory
Pancreatic acinar cells secrete both active enzymes and proenzymes (zymogens):
| Category | Proenzymes (inactive) | Active enzymes |
|---|---|---|
| Proteases | Trypsinogen (anionic, cationic, mesotrypsin), chymotrypsinogen A/B, procarboxypeptidase A (1,2), procarboxypeptidase B (1,2), proelastase, prophospholipase A2, kallireinogen | - |
| Carbohydrases | - | Amylase |
| Lipases | - | Lipase (TG lipase), carboxylesterase, sterol esterase |
| Nucleases | - | DNase, RNase |
- Costanzo Physiology, 7th Ed., Table 8.5; Sleisenger & Fordtran's GI Disease, Box 56.1
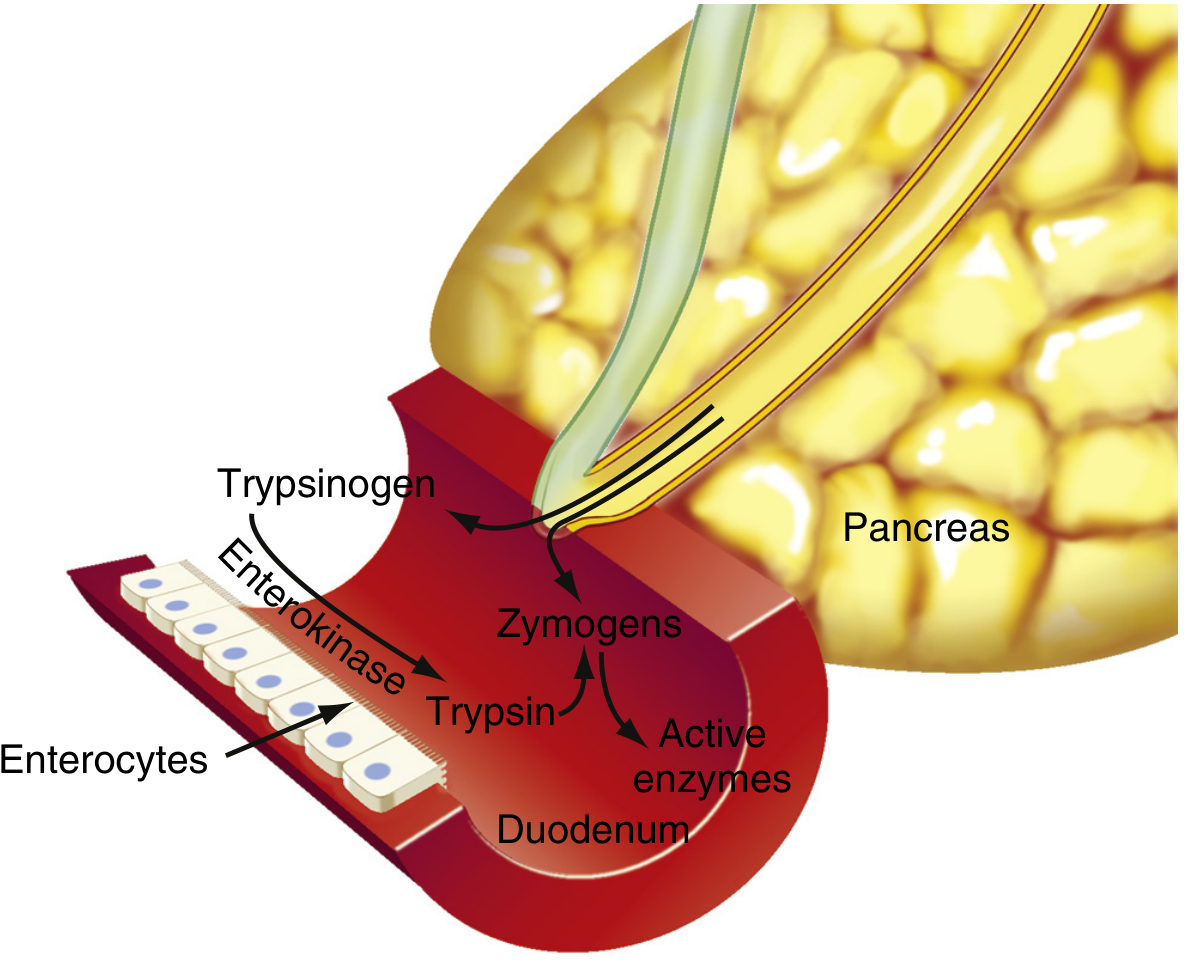
2c. Zymogen Activation Cascade in the Duodenum
The proteolytic enzymes are stored and secreted as inactive precursors - this is essential to prevent self-digestion of the pancreas.
Activation sequence:
- Enterokinase (enteropeptidase) - a brush-border enzyme secreted by duodenal mucosa - cleaves a hexapeptide fragment (Val-Asp-Asp-Asp-Asp-Lys) from trypsinogen, generating active trypsin
- Trypsin also autocatalytically activates more trypsinogen
- Trypsin then activates all other zymogens:
- Chymotrypsinogen → chymotrypsin
- Procarboxypeptidase → carboxypeptidase
- Proelastase → elastase
- Prophospholipase A2 → phospholipase A2
Trypsin inhibitor (PSTI - Pancreatic Secretory Trypsin Inhibitor): A 56-amino acid peptide secreted alongside the zymogens. It inactivates any trypsin formed prematurely within pancreatic ducts by forming a stable complex near the catalytic site. This is the primary defense against autoactivation and acute pancreatitis.
2d. Enzyme Functions
Proteases:
- Trypsin and chymotrypsin - endopeptidases that cleave internal peptide bonds (trypsin at Lys/Arg residues; chymotrypsin at aromatic/bulky hydrophobic residues); produce peptides but not free amino acids
- Carboxypeptidase - exopeptidase that cleaves amino acids from the C-terminus, completing digestion to free amino acids
- Elastase - cleaves at Ala, Gly, Ser residues
Amylase:
- Active enzyme secreted directly (no zymogen form needed - the pancreas contains no starch or glycogen substrate)
- Hydrolyzes 1,4-glycosidic linkages of starch and glycogen at every other junction
- Products: maltose, maltotriose, and alpha-dextrins (containing 1,6-glycosidic branches that amylase cannot cleave - these are finished by brush-border dextrinase)
- Identical enzymatic activity to salivary amylase, but differs in molecular weight, carbohydrate content, and electrophoretic mobility
Lipases:
-
Pancreatic lipase (TG lipase): binds to the oil-water interface of triglyceride droplets; cleaves fatty acids at positions sn-1 and sn-3, releasing 2 fatty acids + 1 monoglyceride (2-monoglyceride). Requires colipase (secreted as procolipase, activated by trypsin) which forms a ternary complex with lipase and bile salts, anchoring lipase to the hydrophobic TG surface. Bile acids emulsify fat to increase surface area.
-
Phospholipase A2: secreted as prophospholipase A2 (activated by trypsin); cleaves the fatty acid ester at carbon 2 of phosphatidylcholine → free fatty acid + lysophosphatidylcholine
-
Carboxylesterase: broad specificity; cleaves cholesterol esters, TGs, diglycerides, monoglycerides, and lipid-soluble vitamin esters. Requires bile salts for full activity.
-
Sleisenger & Fordtran's GI Disease, pp. 1011-1012
3. The Aqueous Bicarbonate Component (Ductal Cells)
3a. Ion Composition and Flow-Rate Dependence
Pancreatic juice is isotonic with plasma at all flow rates. However, bicarbonate and chloride concentrations vary reciprocally with flow rate:
- At low flow (resting): HCO3- ~30 mEq/L, Cl- ~115 mEq/L
- At high flow (secretin-stimulated): HCO3- up to 145 mEq/L, Cl- falls proportionally
- Na+ and K+ remain constant (~plasma levels) at all flow rates
- Total daily volume: ~2.5 L; flow increases from 0.2-0.3 mL/min (resting) to 4.0 mL/min (stimulated)
3b. Cellular Mechanism of HCO3- Secretion
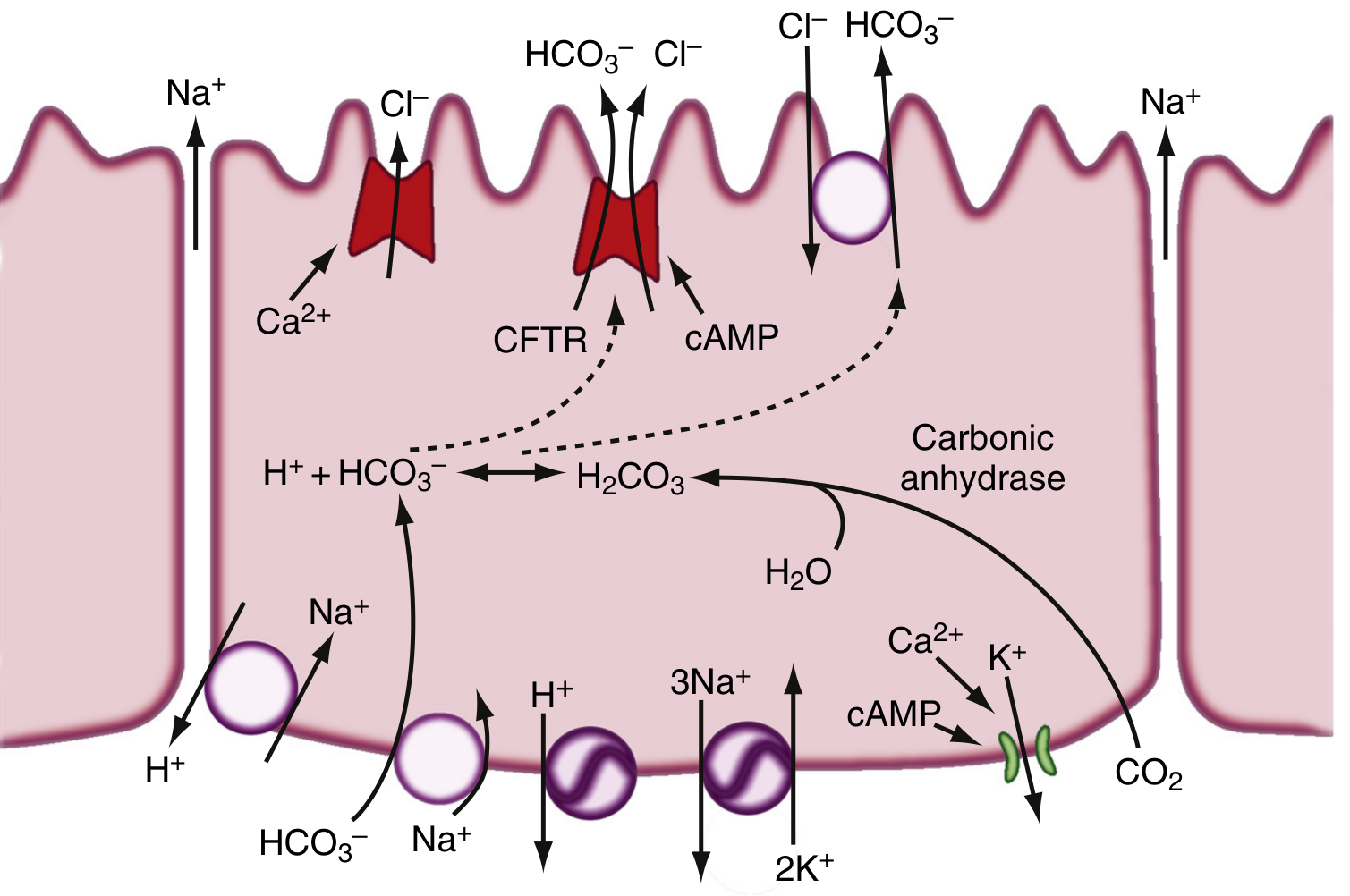
The key steps in bicarbonate generation and secretion:
- CO2 diffuses into the ductal cell from blood (or is generated metabolically)
- Carbonic anhydrase catalyzes: CO2 + H2O → H2CO3 → H+ + HCO3-
- Apical membrane:
- CFTR (Cystic Fibrosis Transmembrane Conductance Regulator) - a cAMP-activated Cl- channel that secretes Cl- into the lumen. CFTR is also directly involved in HCO3- secretion.
- Cl-/HCO3- antiporter (SLC26 family) - exchanges luminal Cl- for intracellular HCO3-, driving net HCO3- secretion into the duct lumen
- Basolateral membrane:
- Na+-K+-ATPase - maintains electrochemical gradients
- Na+/H+ antiporter (NHE) - exports H+ generated by carbonic anhydrase, keeping intracellular pH neutral
- Na+/HCO3- cotransporter - imports HCO3- from blood to replenish intracellular stores
- H+-ATPase - additional H+ exporter
- K+ channel - maintains basolateral potential to drive transport
- Na+ and water follow passively (paracellularly) to maintain isotonicity
Note: In cystic fibrosis, CFTR mutations impair luminal Cl- secretion, reducing the substrate available for the Cl-/HCO3- exchange and thus severely reducing pancreatic HCO3- and fluid output, causing inspissation of ducts and exocrine insufficiency.
- Sleisenger & Fordtran's GI Disease, pp. 1008-1010; Costanzo Physiology, 7th Ed., pp. 372-373
4. Regulation of Pancreatic Secretion
Three phases of secretion occur, similar to gastric secretion:
4a. Phases
| Phase | Stimulus | Mechanism | Contribution |
|---|---|---|---|
| Cephalic | Sight/smell/taste of food | Vagal (ACh) → enzyme secretion | ~20% of enzyme output |
| Gastric | Gastric distension | Vagal reflexes | 5-10% of enzyme output |
| Intestinal | Chyme in duodenum | Secretin + CCK | 70-80% of total secretion |
4b. Key Hormones
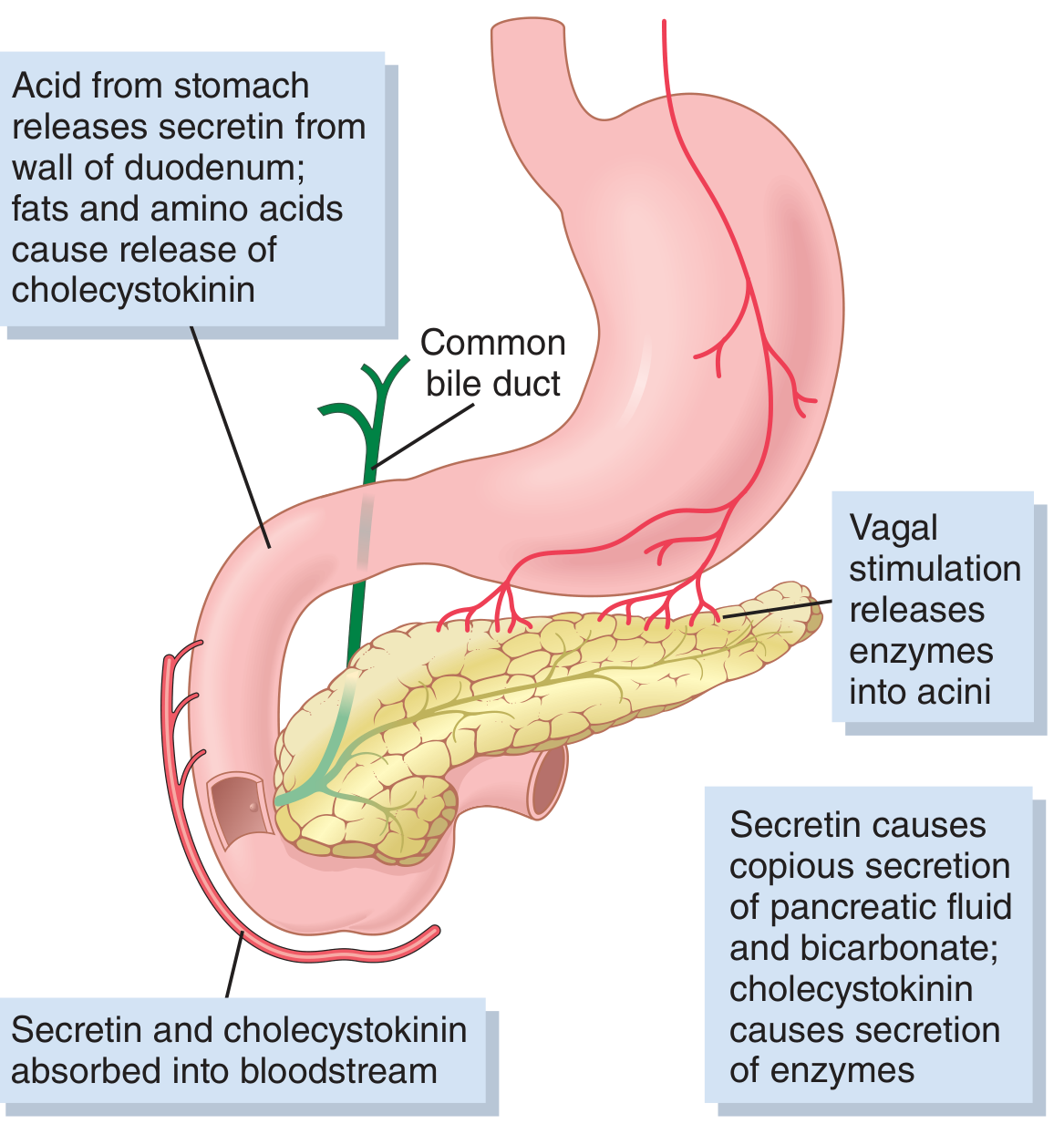
Secretin:
- Released from S cells in duodenal/jejunal mucosa when luminal pH falls below 4.5-5.0 (primary trigger: HCl from stomach)
- 27-amino acid polypeptide; stored as inactive prosecretin
- Acts on ductal cells: binds basolateral receptor → adenylate cyclase → ↑cAMP → PKA activates CFTR and K+ channels → massive HCO3- secretion
- Result: large volume, high HCO3- juice (up to 145 mEq/L); neutralizes duodenal acid
- Neutralization reaction: HCl + NaHCO3 → NaCl + H2CO3 → NaCl + CO2 + H2O
- Antisecretin antibodies reduce meal-stimulated pancreatic HCO3- by 80%, demonstrating its dominance
Cholecystokinin (CCK):
- Released from I cells in duodenal/upper jejunal mucosa; stimulated primarily by proteoses/peptones and long-chain fatty acids in chyme
- 33-amino acid polypeptide
- Acts primarily on acinar cells: binds CCK-A receptor → phospholipase C → IP3 + DAG → ↑intracellular Ca2+ from ER → activation of Ca2+-calmodulin kinase → exocytosis of zymogen granules
- Also signals via protein kinase C (PKC) pathway
- Accounts for 70-80% of total digestive enzyme secretion after a meal
- Has weak effect on duct cell HCO3- secretion
Acetylcholine (ACh):
- Released from parasympathetic (vagal) postganglionic fibers
- Acts on both acinar cells (→ enzyme secretion via Ca2+ pathway, similar to CCK) and ductal cells (→ HCO3- secretion via Ca2+)
- Sympathetic innervation from celiac/superior mesenteric plexus inhibits pancreatic secretion (opposite of salivary glands where both divisions are stimulatory)
Potentiation: When secretin + CCK + ACh act simultaneously, total secretion far exceeds the sum of individual effects - the stimuli multiply (potentiate) each other. This is why all three pathways working together during a meal produce the full secretory response.
Vasoactive Intestinal Peptide (VIP): Similar to secretin in structure; stimulates ductal HCO3- secretion via cAMP.
Gastrin-releasing peptide (GRP/bombesin): Released from enteric neurons; stimulates acinar enzyme secretion.
4c. Interdigestive (Basal) Secretion
Between meals, pancreatic secretion is low (0.2-0.3 mL/min). It shows a cyclic pattern linked to the migrating motor complex (MMC) - secretion peaks during phase III of the MMC.
4d. Feedback Regulation
Trypsin in the duodenal lumen cleaves a CCK-releasing peptide (CCKRP), which is normally secreted by duodenal cells and stimulates CCK release. Once proteins are fully digested, trypsin cleaves CCKRP, reducing CCK stimulation - a negative feedback loop that matches enzyme output to the digestive load.
- Guyton & Hall Medical Physiology; Sleisenger & Fordtran's GI Disease, pp. 1015-1016; Costanzo Physiology, 7th Ed., pp. 373-376
5. Adaptation of Enzyme Synthesis to Diet
The relative synthesis rates of specific enzymes change with chronic dietary composition:
- Carbohydrate-rich diet → ↑ amylase synthesis, ↓ chymotrypsinogen
- Lipid-rich diet → ↑ lipase expression
- Alcohol-rich diet → ↓ amylase expression
- Regulated at both transcriptional and post-transcriptional levels; amylase gene is also regulated by insulin
This allows the exocrine pancreas to tailor its enzyme output to anticipated nutrient loads.
Summary Table
| Component | Cell of Origin | Key Biochemical Process | Primary Regulator |
|---|---|---|---|
| Enzymes (proteases, amylase, lipases) | Acinar cells | RER synthesis → Golgi → zymogen granules → Ca2+-triggered exocytosis | CCK, ACh |
| HCO3- / fluid | Ductal/centroacinar cells | Carbonic anhydrase + CFTR + Cl-/HCO3- antiport | Secretin (cAMP) |
| Trypsin inhibitor (PSTI) | Acinar cells | Co-secreted with zymogens; blocks premature trypsin | - |
| Colipase | Acinar cells | Procolipase activated by trypsin; anchors lipase to fat droplets | CCK |
Sources:
- Costanzo Physiology, 7th Edition - Ch. 8, pp. 371-376
- Guyton and Hall Textbook of Medical Physiology - Ch. 65, pp. 805-807
- Sleisenger and Fordtran's Gastrointestinal and Liver Disease - Ch. 56, pp. 1008-1016
This is a shared conversation. Sign in to Orris to start your own chat.