sensory coding based on guyton
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Reading File
Loading Image
Reading File
I have all the content needed for a complete answer. Here is the comprehensive write-up based entirely on Guyton and Hall.
Sensory Coding - Guyton and Hall
(Guyton and Hall Textbook of Medical Physiology, Chapters 47 & 48)
1. Types of Sensory Receptors
Guyton classifies sensory receptors into five main groups based on the type of stimulus they detect:
| Type | Stimulus | Examples |
|---|---|---|
| Mechanoreceptors | Mechanical compression/stretch | Pacinian corpuscles, Meissner corpuscles, Merkel discs, Ruffini endings, hair receptors, muscle spindles, Golgi tendon organs, baroreceptors |
| Thermoreceptors | Temperature change | Cold receptors, warm receptors |
| Nociceptors | Tissue damage (physical/chemical) | Free nerve endings |
| Electromagnetic receptors | Light energy | Rods and cones of the retina |
| Chemoreceptors | Chemical stimuli | Taste buds, olfactory epithelium, carotid/aortic bodies, hypothalamic glucose/amino acid receptors |
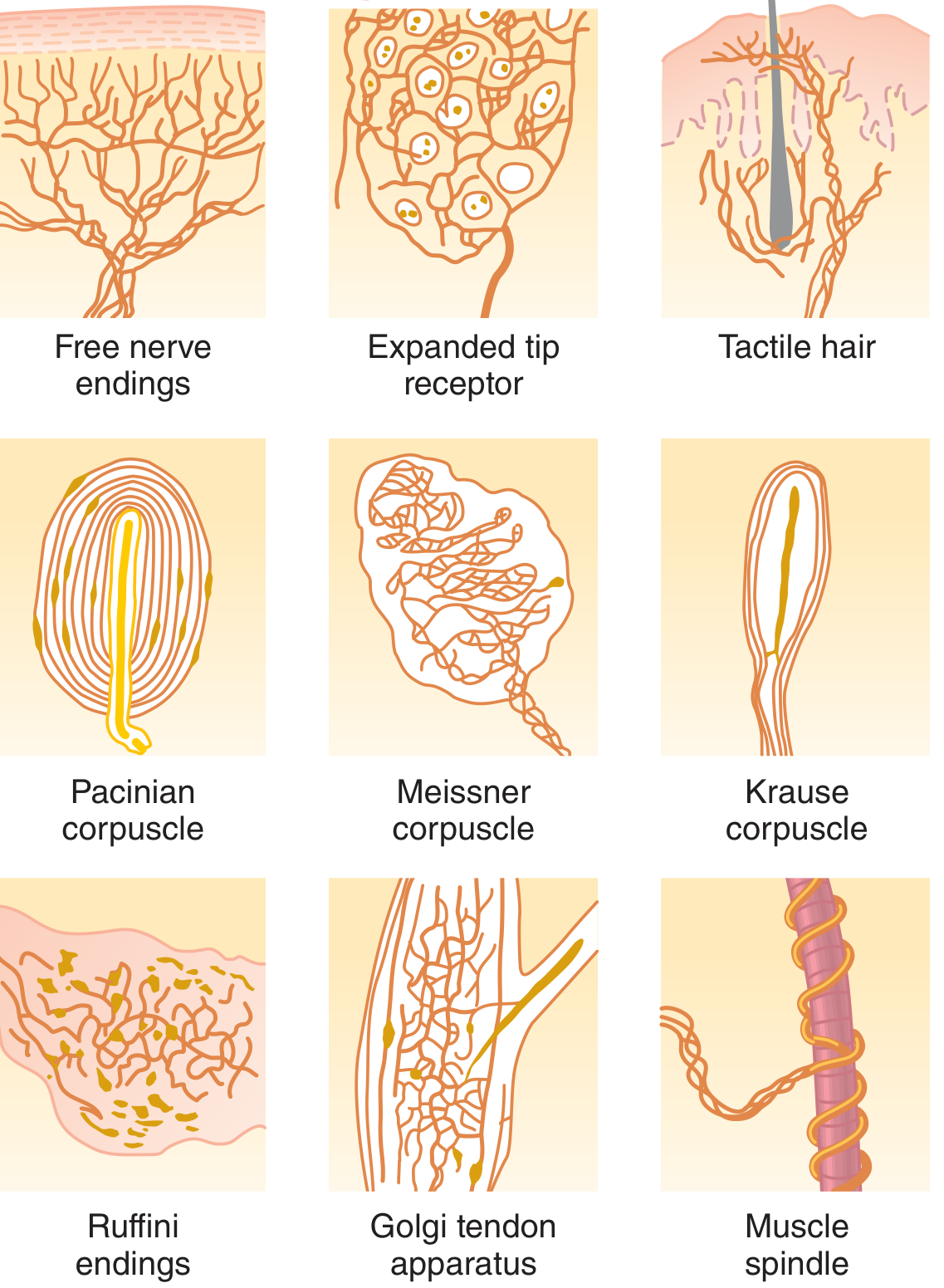
Fig. 47.1 - Types of somatic sensory nerve endings (Guyton & Hall)
2. Differential Sensitivity of Receptors
Each receptor type is highly sensitive to its specific ("adequate") stimulus and almost completely non-responsive to other stimulus types. For example:
- Rods and cones respond to light but not to normal ranges of heat or pressure.
- Osmoreceptors in the supraoptic nucleus detect osmolality changes but not sound.
- Pain receptors are almost never activated by ordinary touch, but are highly active when tissue damage occurs.
This differential sensitivity is the foundation of sensory specificity.
3. Modality of Sensation and Neural Coding
A. Labeled Line Neural Coding
Each modality of sensation (pain, touch, sight, sound, etc.) is transmitted by a specific set of nerve fibers that terminate at specific points in the CNS. The sensation perceived is determined by where in the nervous system the fiber leads, not by the type of stimulus that activates it. This is labeled line coding.
"If a pain fiber is stimulated, the person perceives pain regardless of what type of stimulus excites the fiber - electricity, overheating, crushing, or tissue damage."
Similarly, touch fibers lead to touch areas in the brain, optic fibers lead to visual cortex, and auditory fibers lead to auditory cortex. The label is in the destination, not the message itself.
B. Population Coding
In some cases, a single cell encodes more than one modality, and a combination of signals from many cells is required to determine the final sensation. Color vision is the classic example: each cone type (blue, green, red) is most sensitive to one wavelength but responds to a range. The final color perception emerges from comparing firing rates across all three populations. Even more complex sensations (recognizing an object's shape, color, texture) require activation of large receptor ensembles in parallel, with sequential relay and processing by the CNS.
4. Transduction of Sensory Stimuli into Nerve Impulses
Receptor Potential
All sensory receptors share one common feature: whatever stimulus excites the receptor, its immediate effect is to change the membrane electrical potential of the receptor. This is the receptor potential.
Mechanisms of receptor potential generation:
- Mechanical deformation - stretches receptor membrane, opens stretch-sensitive ion channels
- Chemical application - opens ion channels or activates second messenger systems via membrane receptors
- Temperature change - alters temperature-sensing ion channels and metabotropic receptors
- Electromagnetic radiation - light alters retinal receptor membrane characteristics, changing ion flux
In all cases, the change in membrane permeability allows ions to diffuse across the membrane, changing the transmembrane potential. Maximum receptor potential amplitude is about 100 mV (same as the action potential maximum), reached only at extremely high stimulus intensity.
Receptor Potential Firing Action Potentials
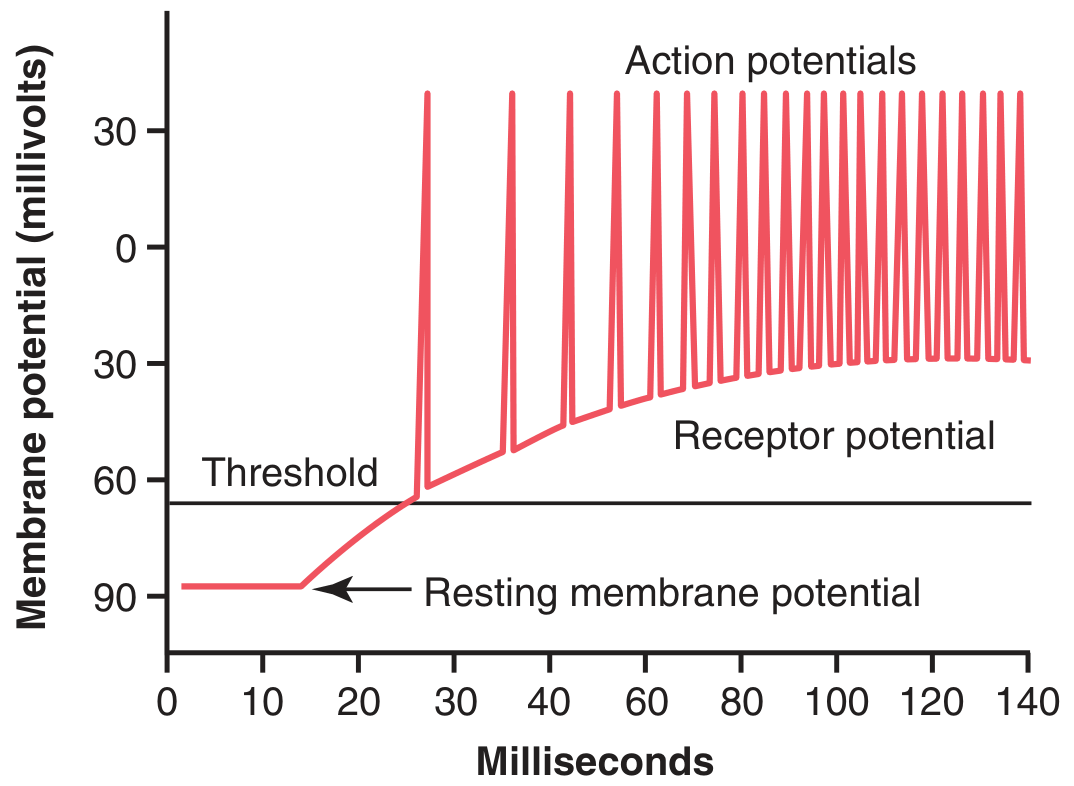
Fig. 47.2 - When the receptor potential rises above threshold, action potentials are generated. The higher the receptor potential above threshold, the greater the action potential frequency (temporal/frequency coding).
Pacinian Corpuscle as a Model
The Pacinian corpuscle illustrates receptor function clearly: compression deforms the central unmyelinated fiber tip, opening ion channels and letting Na+ flow inward, generating the receptor potential. This triggers current flow to the first node of Ranvier, where action potentials are initiated and transmitted centrally.
The receptor potential amplitude increases with stimulus strength but in a non-linear (compressive) fashion - rising steeply at low intensities and less steeply at high intensities. This gives the receptor a wide dynamic range, allowing detection of both very weak and very intense stimuli.
5. Transmission of Signal Intensity - Frequency Coding and Spatial/Temporal Summation
Signal intensity is transmitted to the CNS by two mechanisms:
A. Temporal Summation (Frequency Coding)
As stimulus intensity increases, the receptor potential rises and triggers action potentials at a higher frequency in a single fiber. The CNS interprets higher firing frequency as greater stimulus intensity. This is the primary mechanism of intensity coding.
B. Spatial Summation
Stronger stimuli recruit more parallel nerve fibers simultaneously. Each pain fiber, for example, arborizes into hundreds of free nerve endings covering a receptive field of up to 5 cm diameter. A strong central stimulus activates more fibers than a weak peripheral one. Progressively stronger stimuli spread activation to progressively more fibers in the nerve bundle.
6. Sensory Nerve Fiber Classification
| Classification | Fiber Type | Diameter | Velocity | Function |
|---|---|---|---|---|
| Group Ia | A-alpha | ~17 µm | Fastest (~120 m/s) | Annulospiral endings of muscle spindles |
| Group Ib | A-alpha | ~16 µm | Fast | Golgi tendon organs |
| Group II | A-beta/gamma | ~8 µm | Medium | Discrete tactile receptors, flower-spray spindle endings |
| Group III | A-delta | ~3 µm | Moderate | Temperature, crude touch, pricking pain |
| Group IV | C fibers | 0.5-2 µm | Slowest (~0.5 m/s) | Pain, itch, temperature, crude touch |
The smallest C fibers require about 2 seconds to travel from the big toe to the spinal cord, while the largest A fibers cover that same distance in a fraction of a second.
7. Receptor Adaptation
When a continuous stimulus is applied, receptors respond at a high rate initially, then at a progressively slower rate. This is receptor adaptation.
Rapid adapters (phasic receptors):
- Pacinian corpuscles - adapt to extinction within a few hundredths of a second
- Hair base receptors - adapt within 1 second or so
- These respond maximally to a change in stimulus (onset/offset), not to a sustained one
Slow adapters (tonic receptors):
- Joint capsule and muscle spindle receptors - adapt slowly over minutes to hours
- Carotid/aortic baroreceptors - adapt over ~2 days (some physiologists believe they never fully adapt)
- Chemoreceptors and pain receptors - probably never adapt completely
Mechanism of adaptation:
For the Pacinian corpuscle, the layered capsule itself is responsible for rapid adaptation. When a sustained pressure is applied, the fluid within the capsule redistributes and the central fiber returns to its resting state, removing the receptor potential. If the capsule is stripped away, the naked nerve fiber adapts slowly under the same sustained stimulus - demonstrating it is the capsule mechanics, not the nerve itself, that drives rapid adaptation.
For other receptors (e.g., retinal rods/cones), chemical changes within the receptor itself are responsible for adaptation.
8. Interpretation of Sensory Stimulus Intensity
Weber-Fechner Principle
Stimulus intensity is perceived logarithmically:
Interpreted signal strength = Log(Stimulus) + Constant
A person holding 30 g can just detect 1 g added; holding 300 g requires 10 g added to just detect a change. The ratio (~1:30) stays constant. This is quantitatively accurate for higher intensities of visual, auditory, and cutaneous sensations, but applies poorly to other sensory types.
Power Law (Stevens' Power Law)
A more accurate formula across all sensation types:
Interpreted signal strength = K × (Stimulus - k)^y
Plotted on double-logarithmic coordinates with appropriate constants, this yields a linear relation between interpreted and actual stimulus strength across a large intensity range for almost any sensory modality.
Why is such a wide range needed?
The auditory system handles a dynamic range of over 10 billion-fold in sound intensity (from a whisper to an explosion). The visual system handles 500,000-fold variation in light. If the system were linear, it would either saturate at high intensities or fail to detect weak stimuli. The compressive (logarithmic/power law) coding solves this.
9. Signal Transmission in Nerve Tracts
Sensory signals ascend to the brain primarily through:
- Dorsal column - medial lemniscal pathway: highly myelinated, fast fibers; carries fine touch, vibration, two-point discrimination, proprioception. Can resolve stimuli changing in as little as 1/400 of a second. Vibratory sensation (up to 700 Hz) is carried exclusively in this pathway.
- Anterolateral spinothalamic pathway: carries pain, temperature, crude touch; slower, less precise.
Lateral inhibition (surround inhibition) sharpens spatial discrimination: stimulated neurons in the center of a receptive field inhibit those at the periphery via interneurons, making peaks of excitation stand out clearly from background activity.
10. Cortical Organization of Sensory Coding
The sensory cortex is organized in vertical columns of neurons. Each column detects a different sensory spot on the body and a specific sensory modality. This columnar organization ensures that each modality from each body location is processed in its own dedicated circuit, contributing to the precision of conscious sensory experience.
Summary Table - The "4 Questions" of Sensory Coding
| What is coded? | Mechanism |
|---|---|
| Modality (what type) | Labeled line coding - which pathway/center is activated |
| Intensity (how strong) | Frequency coding (temporal summation) + spatial summation; Weber-Fechner / Power Law for perception |
| Location (where) | Receptive fields, topographic mapping in cortex, lateral inhibition |
| Duration (how long) | Adaptation rate - phasic vs. tonic receptors |
Guyton and Hall Textbook of Medical Physiology, 14th Ed., Chapters 47 (Sensory Receptors, Neuronal Circuits) and 48 (Somatic Sensations), pp. 583-605
This is a shared conversation. Sign in to Orris to start your own chat.