Structure of cell membrane and membrane transport
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Finding Sources
Loading Image
Loading Image
Loading Image
I now have comprehensive content and diagrams from multiple authoritative sources. Let me compose the full detailed answer.
Cell Membrane: Structure and Membrane Transport
Part 1 - Structure of the Cell Membrane
Overview
The cell membrane (plasma membrane) envelops the cell and is a thin, pliable, elastic structure only 7.5 to 10 nanometers thick. Its approximate composition is:
| Component | % |
|---|---|
| Proteins | 55% |
| Phospholipids | 25% |
| Cholesterol | 13% |
| Other lipids | 4% |
| Carbohydrates | 3% |
- Guyton and Hall Textbook of Medical Physiology, p. 32
The Lipid Bilayer
The basic structural framework is a lipid bilayer - a double-layered film of lipids, each layer one molecule thick, continuous over the entire cell surface. It is composed of three main lipid types:
1. Phospholipids (most abundant)
- Each phospholipid has a hydrophilic phosphate head (faces aqueous environments) and hydrophobic fatty acid tails (face inward, away from water).
- The hydrophobic tails are mutually attracted to one another and form the interior of the bilayer; the hydrophilic heads face the intracellular and extracellular water.
- This middle lipid layer is impermeable to water-soluble substances like ions, glucose, and urea. Fat-soluble substances (O₂, CO₂, alcohol) cross with ease.
2. Sphingolipids
- Derived from the amino alcohol sphingosine; also amphipathic.
- Present in small amounts, especially in nerve cells.
- Functions: protection from harmful environmental factors, signal transmission, and adhesion sites for extracellular proteins.
3. Cholesterol
- Highly fat-soluble steroid nucleus; dissolves within the bilayer.
- Controls membrane fluidity and the degree of permeability to water-soluble constituents.
- Guyton and Hall Textbook of Medical Physiology, pp. 32-33
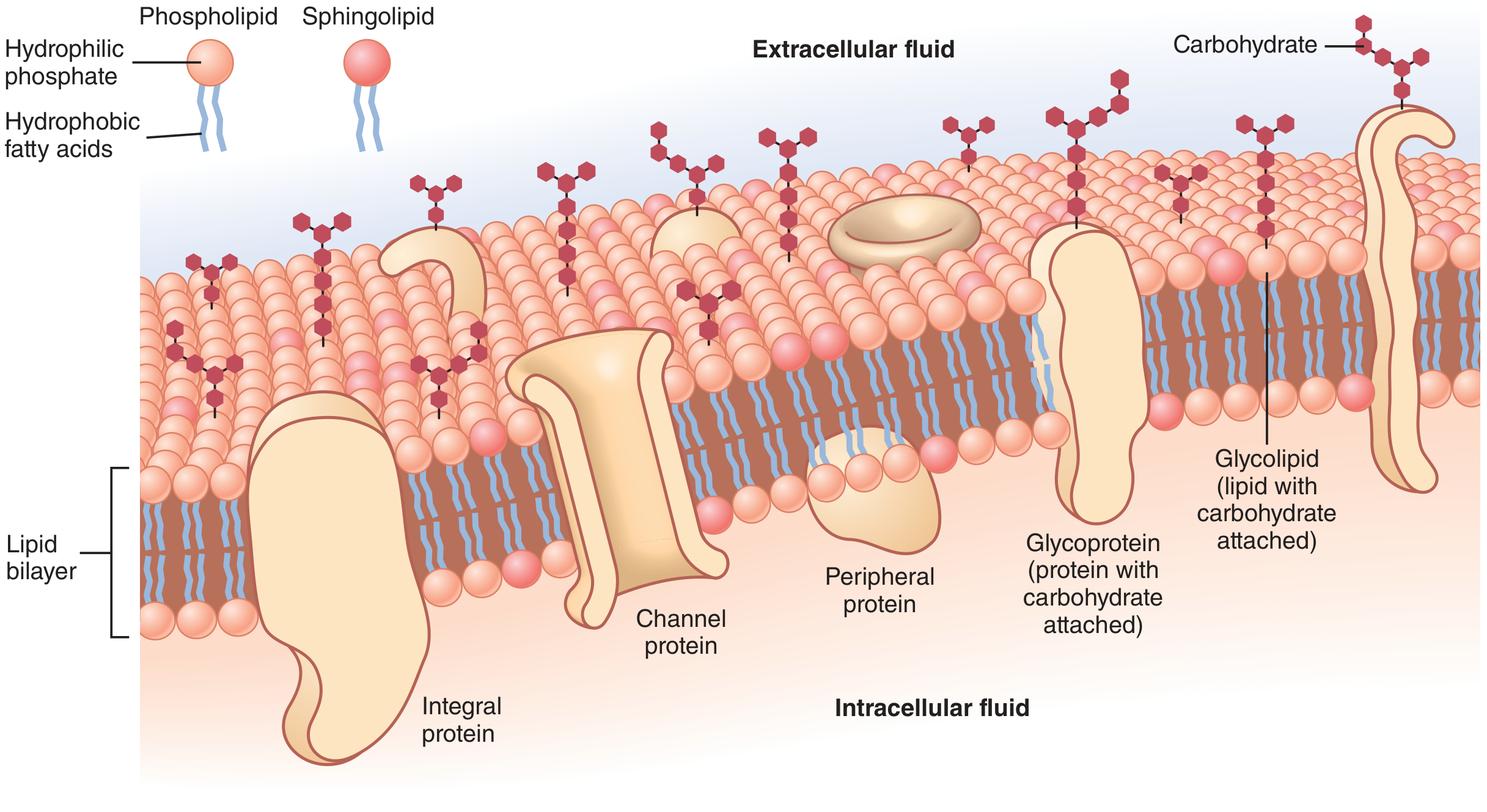
Figure: Structure of the cell membrane (Guyton & Hall). Note the phospholipid and sphingolipid molecules, integral proteins (including channel proteins), peripheral proteins, glycoproteins, and glycolipids.
Membrane Proteins
There are two major classes:
Integral (Intrinsic) Proteins
- Protrude all the way through the membrane (transmembrane proteins).
- Most are glycoproteins.
- Functions:
- Form structural channels (pores) allowing diffusion of water, ions, and small molecules between intracellular and extracellular fluids (with selective properties).
- Act as carrier proteins for substances that cannot penetrate the lipid bilayer (some transport against electrochemical gradients = active transport).
- Act as enzymes.
- Serve as receptors for water-soluble chemicals (e.g., peptide hormones) - ligand binding causes conformational changes that relay signals to the cell interior via second messengers.
Peripheral (Extrinsic) Proteins
- Attached to an integral protein or to one surface only; do not penetrate all the way through.
- Function almost entirely as enzymes or as controllers of transport through membrane pores.
- Guyton and Hall Textbook of Medical Physiology, p. 33
Membrane Carbohydrates - The Glycocalyx
Carbohydrates on the membrane occur almost entirely as:
- Glycoproteins (carbohydrate + protein) - most integral proteins are glycoproteins.
- Glycolipids (carbohydrate + lipid) - about 1/10 of membrane lipids.
- Proteoglycans - mainly carbohydrate chains bound to small protein cores, loosely attached to the outer surface.
Together these form a loose carbohydrate coat on the cell exterior called the glycocalyx. Its functions:
- Negative electrical charge (repels other negatively charged objects, prevents cell aggregation).
- Attaches cells to one another (cell-cell adhesion).
- Acts as receptors for hormones (e.g., insulin) - activates internal protein cascades.
- Enters into immune reactions (self/non-self recognition).
- Guyton and Hall Textbook of Medical Physiology, p. 33
Fluid Mosaic Model
The membrane is not a static structure - proteins "float" within the lipid bilayer like icebergs in a sea of lipids. Membrane fluidity is modulated by:
- Cholesterol content (more cholesterol = less fluid at physiological temperatures)
- Fatty acid saturation (unsaturated fatty acids increase fluidity)
- Temperature
Part 2 - Membrane Transport
All substances crossing the membrane are subject to one of two broad categories: passive transport (no energy required) or active transport (energy required).
A. Passive Transport
Movement of substances down their concentration or electrochemical gradient - no cellular energy expenditure.
1. Simple (Passive) Diffusion
- Small, non-polar (lipophilic) molecules and gases (O₂, CO₂, N₂, steroid hormones, alcohol) diffuse directly through the lipid bilayer.
- Driven purely by the concentration gradient and kinetic energy of molecules.
- Water crosses very slowly by this route; it mainly uses aquaporins (see below).
- Example: Exchange of O₂ and CO₂ between blood and tissues.
- Junqueira's Basic Histology, p. 79
2. Facilitated Diffusion
Still moves down the concentration gradient but requires membrane transport proteins. Two subtypes:
a) Channel-mediated transport
- Multipass transmembrane proteins form hydrophilic pores.
- Contain a pore domain that partially penetrates the bilayer and acts as an ion-selectivity filter (3D structure determines which ions pass).
- Ion-selective and regulated:
- Voltage-gated channels - opened/closed by changes in membrane potential (e.g., Na⁺ channels in neurons during action potentials).
- Ligand-gated channels - opened by binding of a neurotransmitter/chemical (e.g., acetylcholine receptor at neuromuscular junctions).
- Mechanically gated channels - opened by mechanical stress (e.g., in inner ear hair cells).
- Aquaporins - specialized channel proteins for water transport.
b) Carrier-mediated transport
- Carrier proteins bind the substrate specifically, undergo conformational changes, and release the molecule on the other side.
- Highly selective - often transport only one type of molecule.
- Example: Glucose carriers (GLUT transporters).
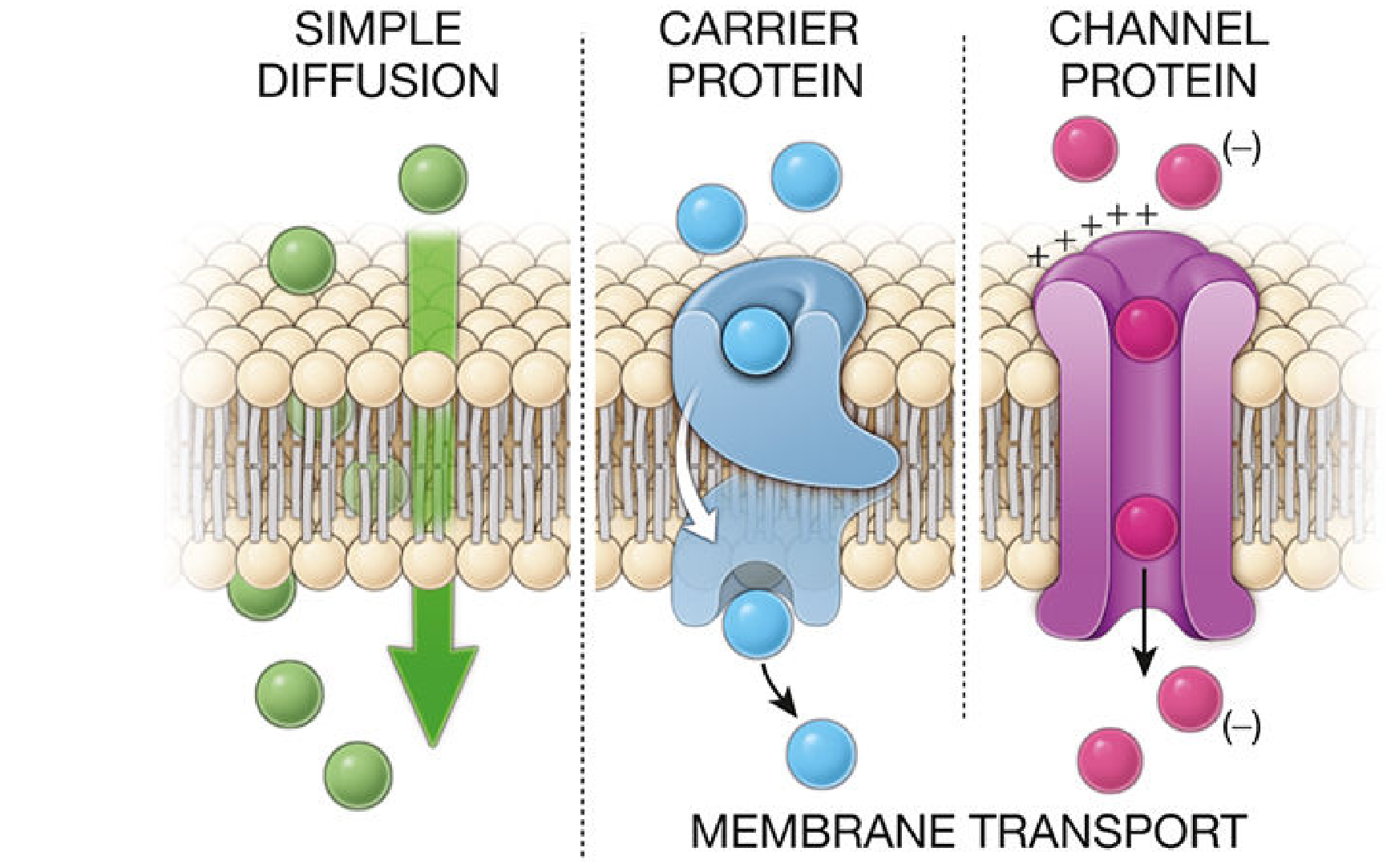
Figure: Simple diffusion (left), carrier-mediated transport (center), and channel-mediated transport (right). (Histology - A Text and Atlas, p. 143)
B. Active Transport
Movement of substances against their concentration or electrochemical gradient; requires ATP expenditure.
Primary Active Transport
Directly coupled to ATP hydrolysis via membrane pump proteins (ATPases).
- Na⁺/K⁺-ATPase (sodium-potassium pump): Pumps 3 Na⁺ out and 2 K⁺ into the cell per ATP molecule. Fundamental for maintaining resting membrane potential.
- H⁺ pump (proton pump): Moves H⁺ against its gradient (e.g., in gastric parietal cells for acid secretion, in mitochondria).
- Ca²⁺-ATPase: Pumps Ca²⁺ out of the cytoplasm.
Secondary Active Transport (Co-transport)
Uses the electrochemical gradient established by primary active transport (usually the Na⁺ gradient) to drive uphill transport of another molecule. Two forms:
- Symport (co-transport): Both Na⁺ and the molecule move in the same direction (e.g., Na⁺/glucose co-transporter in intestinal epithelium).
- Antiport (counter-transport/exchange): Na⁺ and the molecule move in opposite directions (e.g., Na⁺/H⁺ exchanger).
Transporter Superfamilies
As noted in Goodman & Gilman's (the pharmacology reference for drug transport):
- ABC (ATP-Binding Cassette) transporters: Primary active transporters using ATP hydrolysis. ~2000 transporter genes = ~7% of the human genome. Key examples: P-glycoprotein (Pgp/ABCB1, MDR1) - effluxes drugs; CFTR (ABCC7) - chloride channel mutated in cystic fibrosis.
- SLC (Solute Carrier) superfamily: ~65 families with ~460 transporters; includes facilitated and secondary active transporters. Critical for drug absorption and elimination.
- Goodman & Gilman's The Pharmacological Basis of Therapeutics, p. 320
C. Vesicular Transport
Used for large molecules (proteins, polysaccharides, lipid complexes) that cannot cross the membrane via channels or carriers. Involves configurational changes in the plasma membrane forming or fusing vesicles.
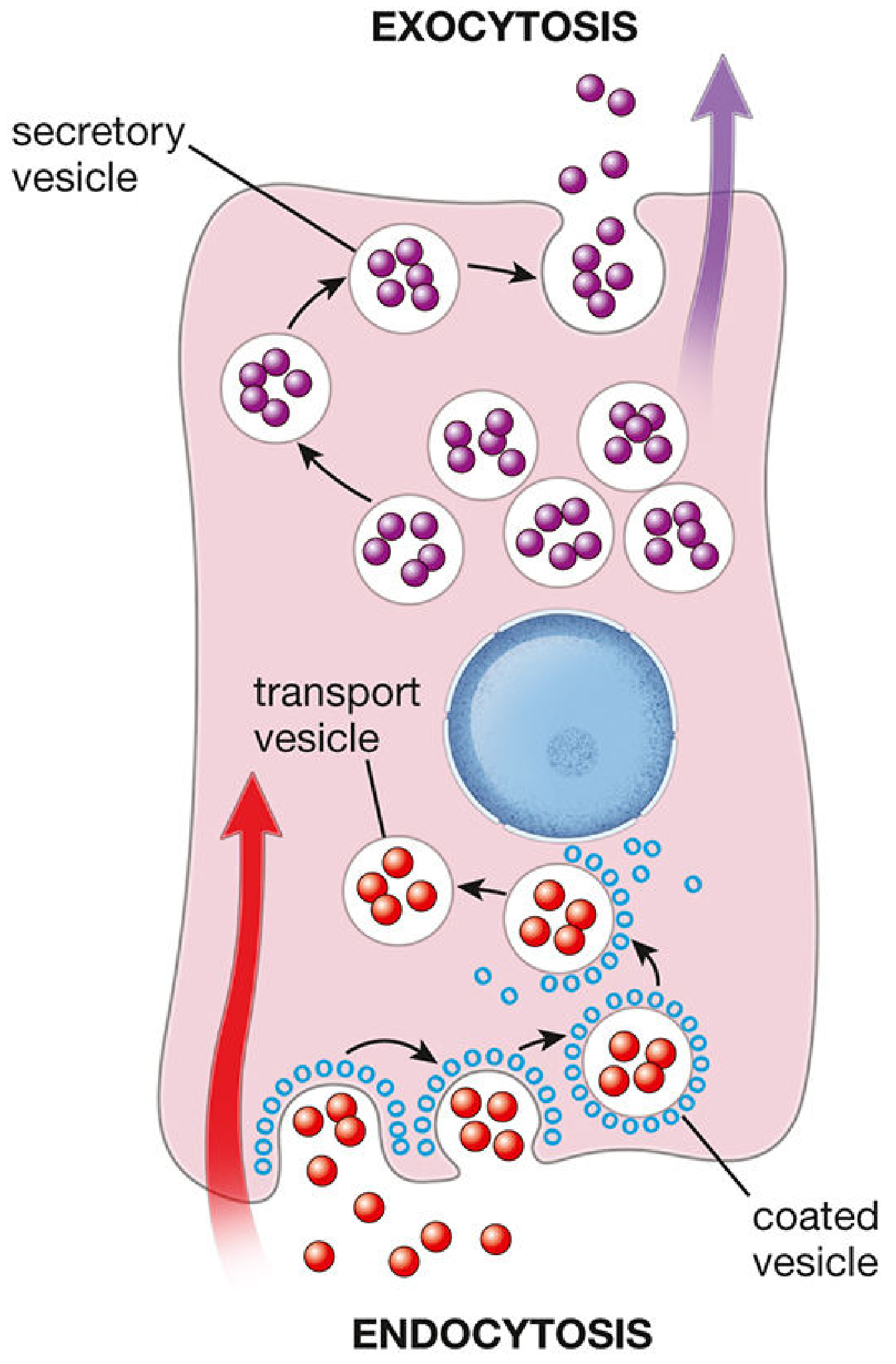
Figure: Endocytosis and exocytosis - the two major forms of vesicular transport. (Histology - A Text and Atlas, p. 144)
Endocytosis (substances enter the cell)
Three main mechanisms:
- Phagocytosis ("cell eating") - engulfment of large particles (bacteria, debris) by pseudopod extension; primarily in macrophages and neutrophils.
- Pinocytosis ("cell drinking") - non-specific uptake of extracellular fluid and small solutes in small vesicles.
- Receptor-mediated endocytosis - the most selective form; specific ligands bind to membrane receptors concentrated in clathrin-coated pits, which invaginate to form coated vesicles. Example: LDL cholesterol uptake via LDL receptors.
Exocytosis (substances leave the cell)
- Intracellular vesicles (from Golgi apparatus) fuse with the plasma membrane, releasing cargo into the extracellular space.
- Primary secretory pathway for hormones, neurotransmitters, enzymes, and antibodies.
- Mediated by SNARE proteins that facilitate vesicle-membrane fusion.
- Exocytosis and endocytosis are coupled - blocking one (e.g., tetanus/botulinum toxins block exocytosis via SNARE proteins) also blocks the other; this prevents secretory cells from swelling or shrinking.
- Histology: A Text and Atlas with Correlated Cell and Molecular Biology, pp. 143-145
Summary Table: Membrane Transport Mechanisms
| Mechanism | Energy | Protein Required | Direction | Examples |
|---|---|---|---|---|
| Simple diffusion | No | No | Down gradient | O₂, CO₂, steroid hormones |
| Channel-mediated | No | Yes (channel) | Down gradient | Na⁺, K⁺, Ca²⁺, water (aquaporins) |
| Carrier-mediated (facilitated) | No | Yes (carrier) | Down gradient | Glucose (GLUT transporters) |
| Primary active transport | Yes (ATP) | Yes (pump) | Against gradient | Na⁺/K⁺ pump, H⁺ pump |
| Secondary active transport | Yes (indirect) | Yes (co-transporter) | Against gradient for one solute | Na⁺/glucose (symport), Na⁺/H⁺ (antiport) |
| Endocytosis | Yes | Membrane remodeling | Into cell | LDL, bacteria, large proteins |
| Exocytosis | Yes | SNARE proteins | Out of cell | Neurotransmitters, hormones |
- Junqueira's Basic Histology, p. 80; Histology: A Text and Atlas, pp. 143-145; Guyton and Hall, pp. 32-34
This is a shared conversation. Sign in to Orris to start your own chat.