excitation contraction coupling physiology and explain for 10 marks 0
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
I now have all the information needed from Guyton & Hall and Costanzo Physiology to write a thorough 10-mark answer. Let me compile it.
Excitation-Contraction Coupling (E-C Coupling)
Definition: Excitation-contraction (E-C) coupling refers to the sequence of events by which an action potential in the muscle fiber membrane is converted into mechanical contraction of the myofibrils. It bridges the electrical event (action potential) and the mechanical event (cross-bridge cycling and shortening). - Guyton & Hall Textbook of Medical Physiology
I. Structural Basis
Before understanding the process, three key structural components must be recognized:
| Structure | Description |
|---|---|
| Sarcolemma | Surface membrane of the muscle fiber; conducts action potentials |
| T Tubules (Transverse tubules) | Invaginations of the sarcolemma that penetrate deep into the fiber; contain extracellular fluid; transmit action potentials to the interior |
| Sarcoplasmic Reticulum (SR) | Specialized endoplasmic reticulum; stores and releases Ca²⁺; composed of terminal cisternae (flanking T tubules) and longitudinal tubules |
The triad consists of one T tubule flanked by two terminal cisternae of the SR. In skeletal muscle, triads are located at the A-I band junction (two per sarcomere in mammalian skeletal muscle).
Fig. 7.5 - T Tubule-Sarcoplasmic Reticulum System:
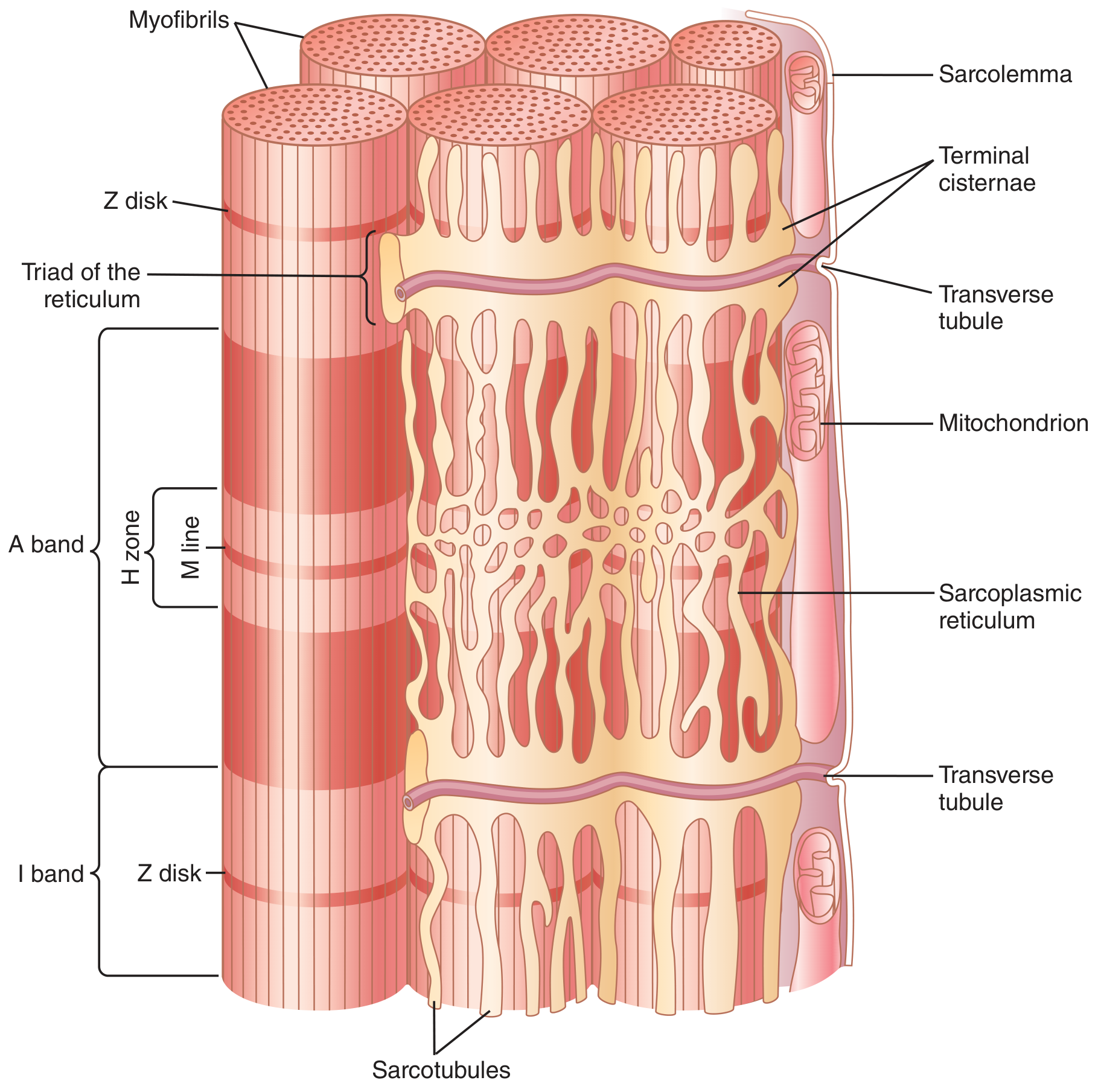
II. E-C Coupling in Skeletal Muscle (Step-by-Step)
Step 1: Action Potential in the Sarcolemma
A motor nerve action potential reaches the neuromuscular junction, releases ACh, which binds nicotinic receptors on the motor end plate, generating an end-plate potential. This triggers an action potential that spreads across the sarcolemma of the entire muscle fiber at 3-5 m/sec.
Step 2: Propagation Down T Tubules
The action potential spreads along the sarcolemma and dips into the T tubule system, penetrating to the deep interior of the muscle fiber. Since the T tubules are open to the extracellular space, the electrical signal reaches the vicinity of every myofibril. This solves the problem of the fiber being too large for surface depolarization to activate the core efficiently.
Step 3: Activation of Dihydropyridine Receptors (DHPR)
At the triad junction, T tubule depolarization is sensed by voltage-sensitive dihydropyridine receptors (DHPR) - L-type Ca²⁺ channels located in the T tubule membrane. In skeletal muscle, DHPRs act primarily as voltage sensors, not as Ca²⁺ channels (unlike cardiac muscle).
Step 4: Mechanical Coupling to Ryanodine Receptors (RyR1)
The activated DHPR is physically coupled (via cytoplasmic loops) to Ryanodine Receptor type 1 (RyR1) channels on the terminal cisternae of the SR. This direct mechanical/conformational coupling causes RyR1 to open.
Step 5: Ca²⁺ Release from the SR
Opening of RyR1 channels releases a large flood of Ca²⁺ from the SR into the sarcoplasm. The resting cytosolic Ca²⁺ concentration is very low (< 10⁻⁷ M). Upon full activation, it rises up to 2 × 10⁻⁴ M - a ~500-fold increase - which is approximately 10 times the level needed for maximum contraction. - Guyton & Hall
Step 6: Ca²⁺ Binds Troponin C
Ca²⁺ diffuses among the myofibrils and binds to troponin C (TnC), the Ca²⁺-binding subunit of the troponin complex on the thin filament. Troponin has three subunits:
- TnC - binds Ca²⁺
- TnI - inhibitory subunit (inhibits actin-myosin interaction at rest)
- TnT - anchors troponin to tropomyosin
Step 7: Removal of Tropomyosin Block
When Ca²⁺ binds TnC, a conformational change occurs in the troponin complex. TnI moves away from actin, and tropomyosin shifts from its blocking position (over the myosin-binding sites on actin) to expose those sites.
Step 8: Cross-Bridge Formation and Contraction
With the myosin-binding sites on actin now exposed:
- Myosin heads bind actin - forming cross-bridges
- The power stroke occurs (ADP + Pᵢ released) - the myosin head pivots, pulling the thin filament toward the M line
- ATP binds myosin head - cross-bridge detaches
- ATP is hydrolyzed to ADP + Pᵢ, re-cocking the myosin head
- Cycle repeats as long as Ca²⁺ is elevated - sliding filament mechanism
Step 9: Relaxation - SERCA Pump
When the action potential ceases, Ca²⁺ is actively pumped back into the SR by the SERCA pump (Sarcoplasmic Reticulum Ca²⁺-ATPase). This pump can concentrate Ca²⁺ up to 10,000-fold inside the SR. Ca²⁺ is stored bound to calsequestrin (binds up to 40 Ca²⁺ per molecule). As cytosolic Ca²⁺ falls, it dissociates from TnC, tropomyosin returns to its blocking position, and the muscle relaxes.
III. Key Differences in Cardiac Muscle E-C Coupling
In cardiac muscle, the mechanism differs in one critical way - Calcium-Induced Calcium Release (CICR):
| Feature | Skeletal Muscle | Cardiac Muscle |
|---|---|---|
| DHPR role | Voltage sensor only (mechanical coupling to RyR) | Also functions as L-type Ca²⁺ channel |
| Ca²⁺ source | Almost entirely from SR (DHPR do not need to conduct Ca²⁺) | From ECF via L-type channels + SR (CICR) |
| RyR type | RyR1 | RyR2 |
| Trigger | Mechanical conformational change | Small trigger Ca²⁺ influx from ECF |
| Extracellular Ca²⁺ dependence | Low | High (heart stops in Ca²⁺-free solution) |
| T tubule diameter | Smaller | 5x larger, rich in mucopolysaccharides storing Ca²⁺ |
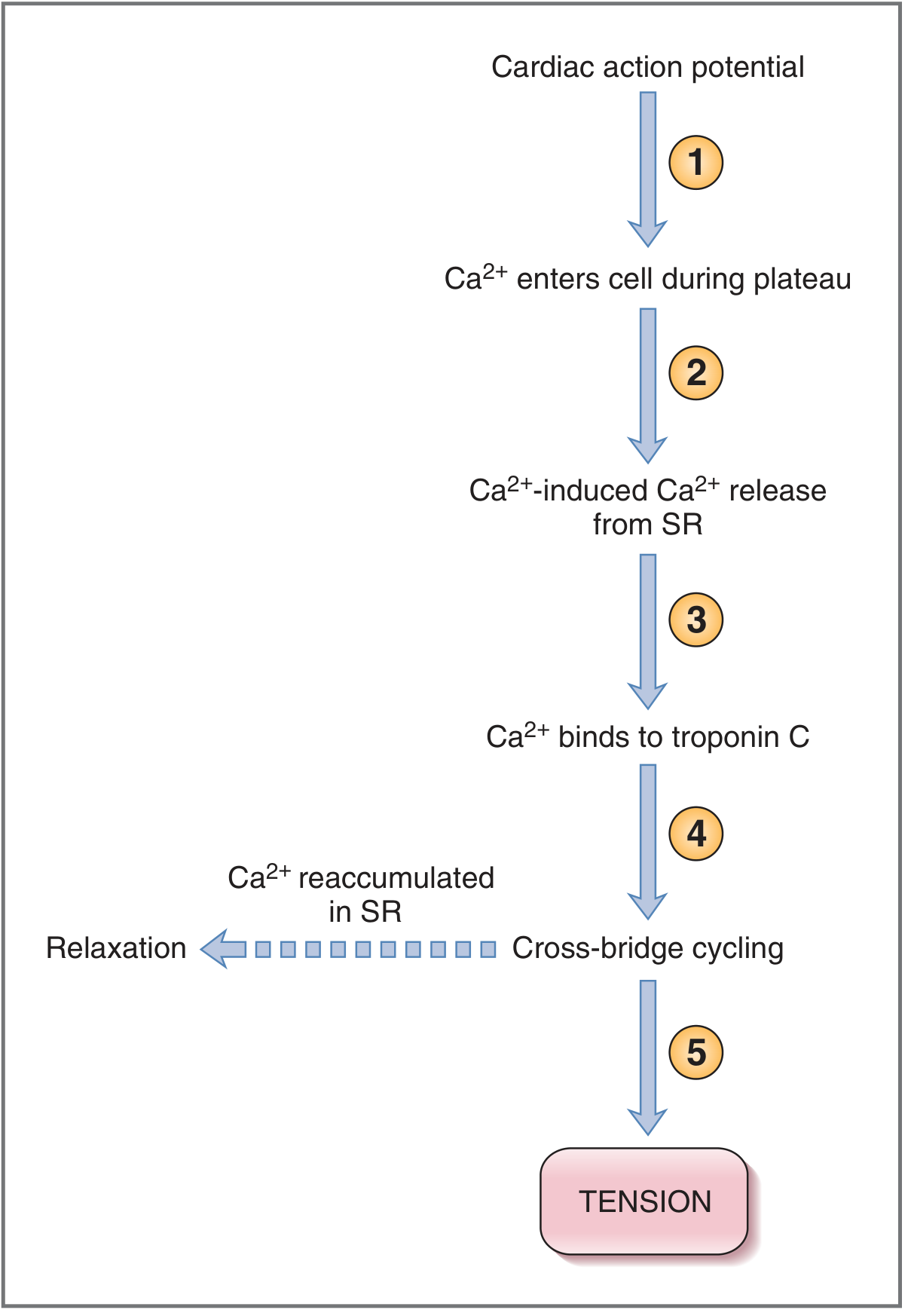
In cardiac muscle: The action potential plateau (Phase 2) opens L-type Ca²⁺ channels in the T tubule, allowing a small amount of Ca²⁺ to enter the cell. This "trigger Ca²⁺" activates RyR2 on the SR, causing a much larger release of Ca²⁺ from SR stores - this amplification mechanism is called Ca²⁺-Induced Ca²⁺ Release (CICR). - Costanzo Physiology 7e
Fig. 9.7 - Cardiac E-C Coupling and Relaxation:
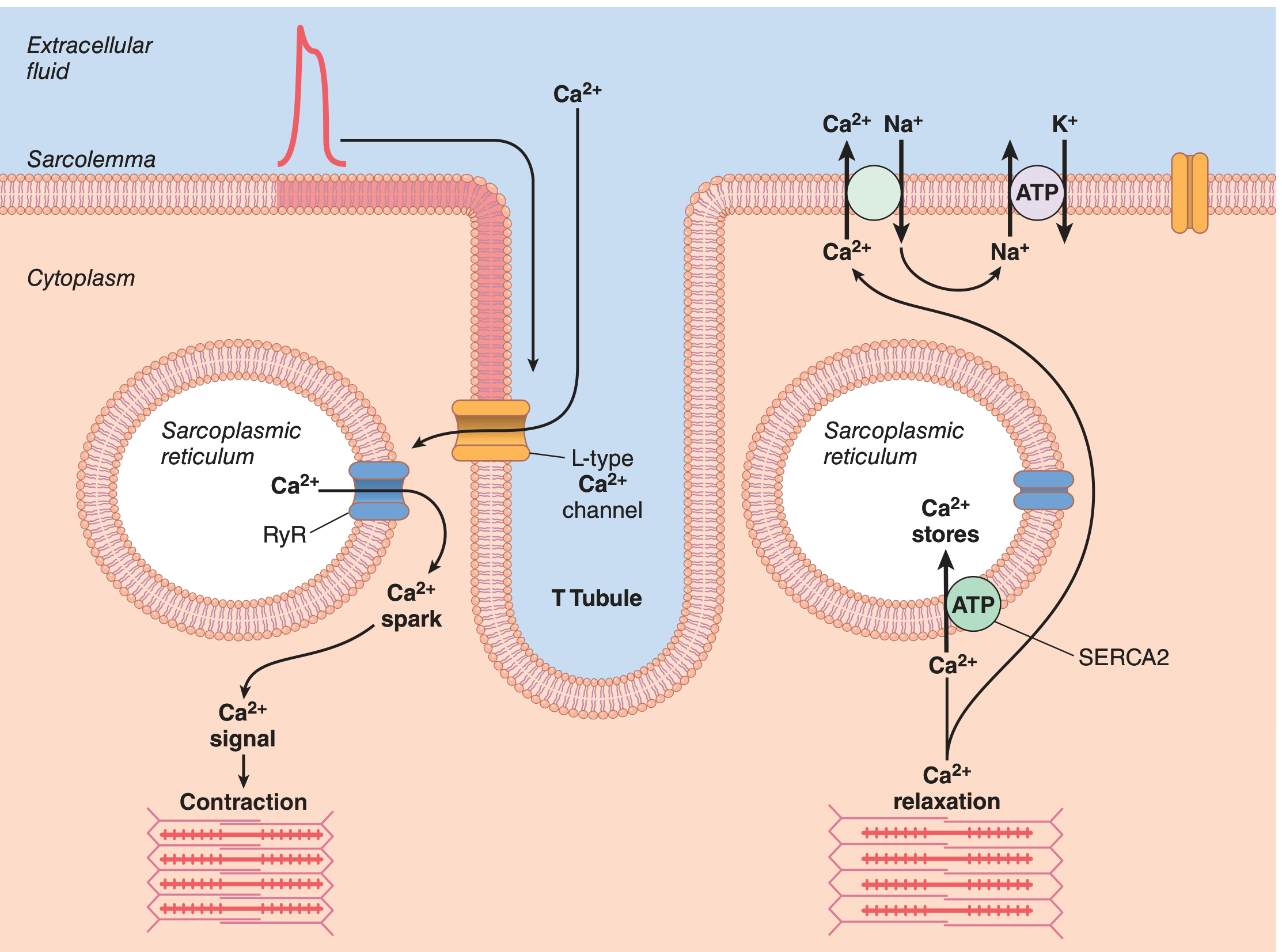
Cardiac E-C Coupling Steps (Costanzo):
Relaxation in cardiac muscle occurs when Ca²⁺ is:
- Reaccumulated into the SR by SERCA2 (using ATP)
- Extruded from the cell by sarcolemmal Ca²⁺-ATPase
- Extruded by NCX (Na⁺-Ca²⁺ exchanger) - uses the inward Na⁺ gradient to pump Ca²⁺ out
IV. Summary Flow Chart
Motor nerve AP
↓
ACh released at NMJ
↓
End-plate potential → Muscle AP (sarcolemma)
↓
AP propagates along T tubules (deep into fiber)
↓
DHPR activated by depolarization
↓
RyR (SR) opens → Ca²⁺ floods sarcoplasm
↓
Ca²⁺ binds Troponin C
↓
Tropomyosin shifts → actin-myosin binding sites exposed
↓
Cross-bridge formation + Power stroke (ATP-driven)
↓
CONTRACTION (sliding filament theory)
↓
SERCA pump active → Ca²⁺ returns to SR
↓
RELAXATION
V. Key Points to Remember (Exam Focus)
- E-C coupling is Ca²⁺-mediated - the entire process pivots on cytosolic Ca²⁺ concentration
- DHPR = voltage sensor in T tubule; RyR = Ca²⁺ release channel in SR
- Skeletal: DHPR physically/mechanically coupled to RyR1 (no Ca²⁺ influx needed from ECF)
- Cardiac: CICR - small Ca²⁺ trigger from ECF (via L-type channel) triggers large Ca²⁺ release from SR via RyR2
- Resting Ca²⁺ < 10⁻⁷ M → during full activation → up to 2 × 10⁻⁴ M (500-fold rise)
- SERCA pump (uses ATP) is responsible for relaxation by pumping Ca²⁺ back into SR
- Cardiac muscle depends heavily on extracellular Ca²⁺ (removing it stops the heart); skeletal muscle does not
- Calsequestrin in the SR lumen stores Ca²⁺ bound to protein (up to 40 Ca²⁺ per molecule)
Sources: Guyton and Hall Textbook of Medical Physiology (14e), pp. 109-110, 125; Costanzo Physiology (7e), pp. 34-38, 150
This is a shared conversation. Sign in to Orris to start your own chat.