Scapula
Reading File
Finding Sources
Searching Images
scapula anatomy bones landmarks
Reading File
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Reading File
Reading File
Loading Image
Loading Image
Finding Sources
Reading File
Reading File
Now I have comprehensive material. Let me compile the full answer.
The Scapula
The scapula is a large, flat, triangular bone on the posterolateral aspect of the thoracic wall, overlying ribs 2–7. It connects to the humerus via the glenohumeral joint and to the axial skeleton via the clavicle at the acromioclavicular joint.
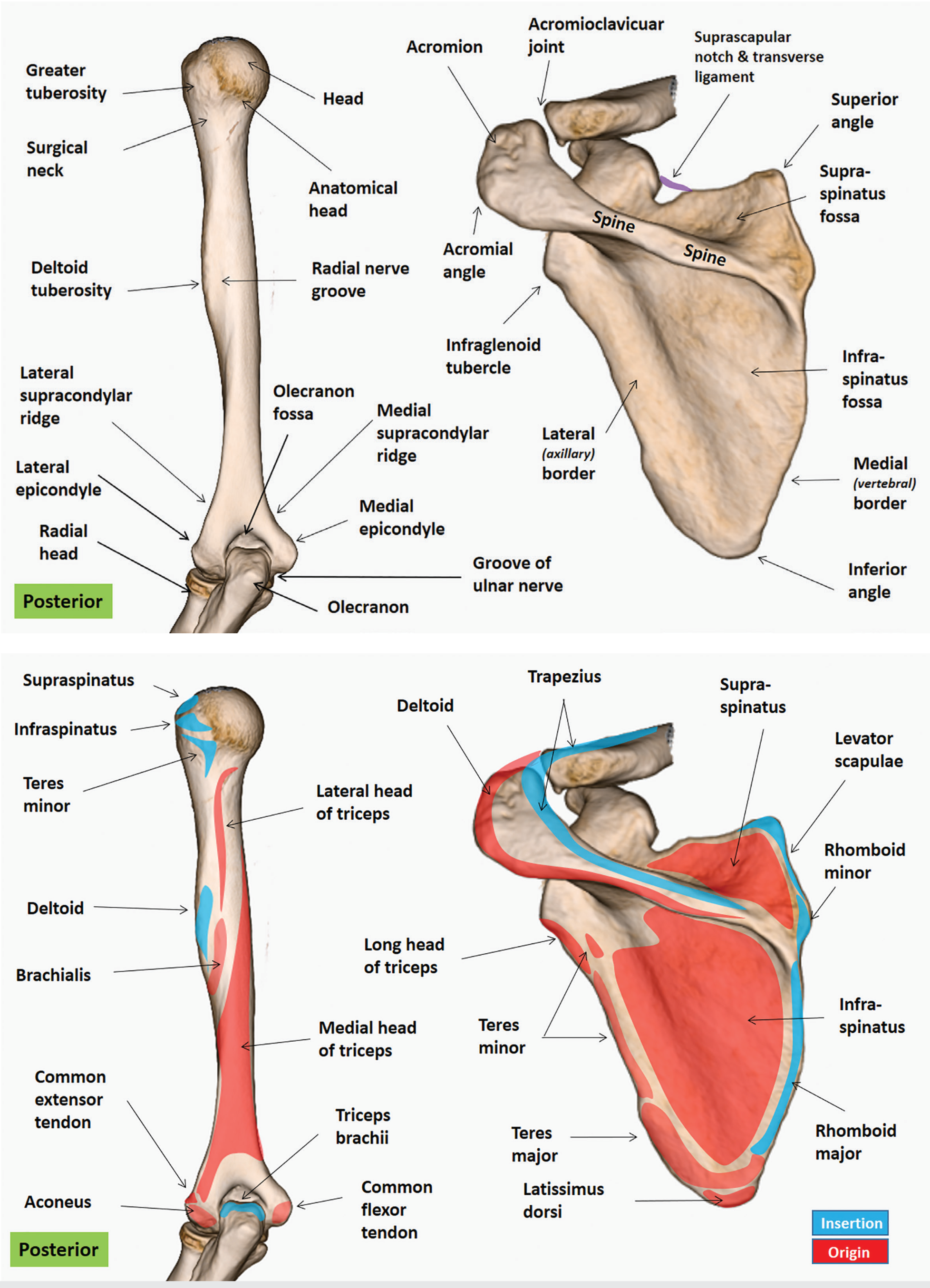
Bony Features
Surfaces (2)
| Surface | Key Feature |
|---|---|
| Costal (ventral/anterior) | Subscapular fossa — shallow concavity where subscapularis attaches; serratus anterior attaches along medial edge |
| Dorsal (posterior) | Divided by the spine of the scapula into supraspinous fossa (supraspinatus origin) above and infraspinous fossa (infraspinatus origin, teres minor/major) below; the two fossae converge at the spinoglenoid notch |
Borders (3)
- Superior border — concave; has the suprascapular notch (covered by the transverse scapular ligament); the inferior belly of omohyoid originates near its lateral end
- Medial (vertebral) border — slightly convex, thickened; site of serratus anterior insertion along its anterior costal surface
- Lateral (axillary) border — thickened; contains the infraglenoid tubercle inferiorly (long head of triceps origin)
Angles (3)
- Superior angle — levator scapulae inserts here
- Inferior angle — the most mobile point; marks the level of T7–T8; serratus anterior holds it against the thorax
- Lateral angle — bears the glenoid cavity (see below)
Processes (3)
- Spine — triangular ridge on dorsal surface; trapezius attaches to its superior surface, deltoid to its inferior surface
- Acromion — continuous with the lateral spine, overhangs the glenohumeral joint; articulates with the clavicle; may fail to fuse (os acromiale, 7–15%)
- Coracoid process — projects anterosuperiorly; coracobrachialis + short head of biceps brachii arise from its tip; pectoralis minor from its medial aspect
Glenoid Cavity
- Shallow, comma-shaped socket at the lateral angle
- ~6–8 cm² in adults, retroverted ~4–8°
- Supraglenoid tubercle — long head of biceps brachii (intra-articular tendon)
- Infraglenoid tubercle — long head of triceps brachii (extra-articular)
Muscle Attachments (17 total)
The scapula has 17 muscular attachments divided into axial stabilizers and scapulohumeral movers.
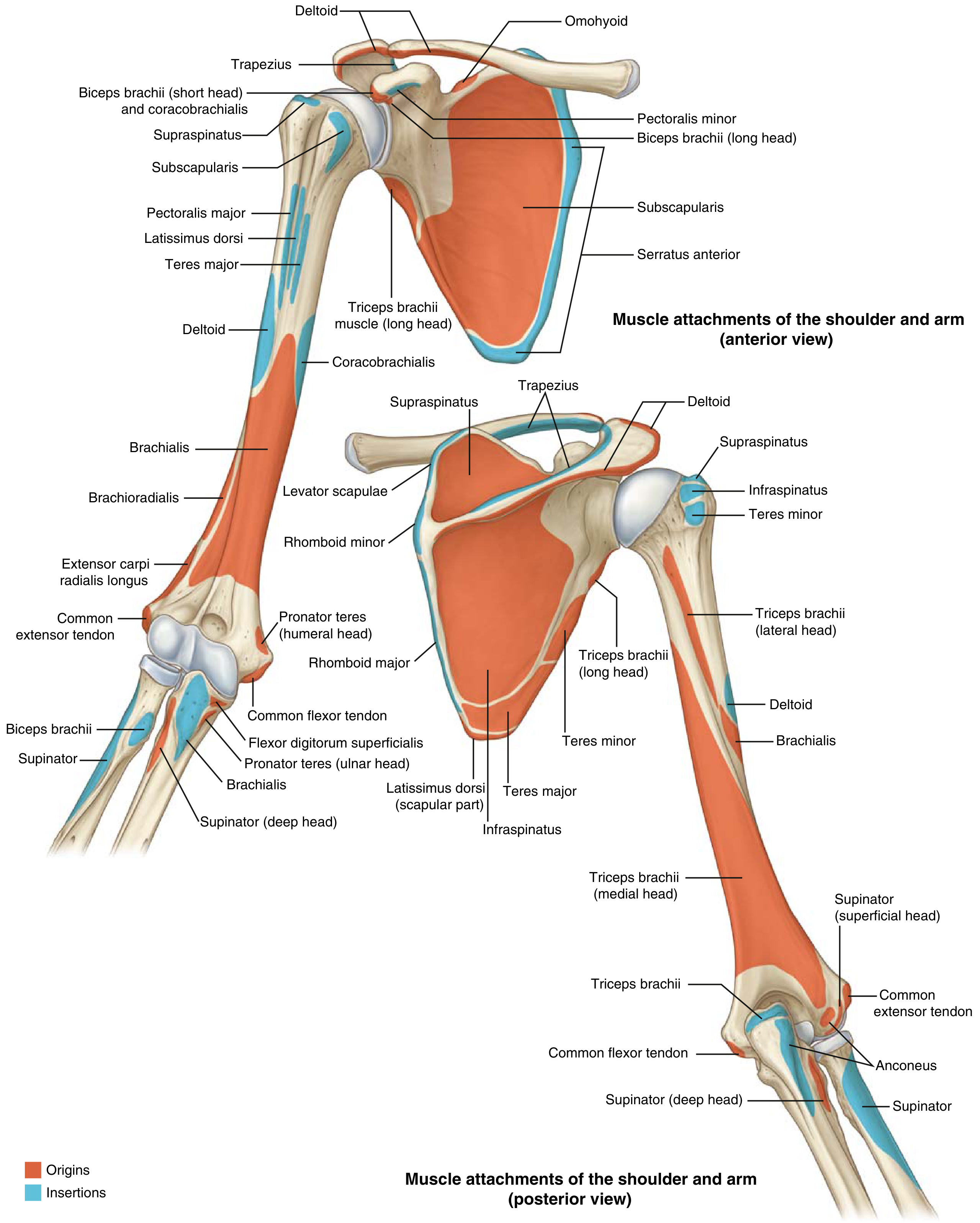
Axial Stabilizers (attach scapula to spine/chest wall)
| Muscle | Origin → Insertion on Scapula | Action |
|---|---|---|
| Trapezius (descending) | Occiput/nuchal lig → lateral clavicle & spine | Elevates, upwardly rotates scapula |
| Trapezius (horizontal) | C7–T3 → acromion & lateral spine | Retracts scapula |
| Trapezius (ascending) | T3–T12 → medial spine | Retracts + depresses |
| Levator scapulae | C1–C4 transverse processes → superior angle | Elevates + rotates |
| Rhomboid minor/major | C6–C7 / T1–T4 spinous processes → medial border | Retraction + elevation |
| Serratus anterior | Ribs 1–10 → anterior surface of medial border | Protraction; upward rotation; holds medial border to thorax |
| Pectoralis minor | Ribs 3–5 → coracoid process | Protraction + depression |
Scapulohumeral Muscles (rotator cuff + others)
| Muscle | Origin | Action |
|---|---|---|
| Supraspinatus | Supraspinous fossa | Initiates abduction, assists ER |
| Infraspinatus | Infraspinous fossa | External rotation |
| Teres minor | Dorsolateral border | External rotation |
| Teres major | Inferior angle (posterior) | IR, adduction, extension |
| Subscapularis | Subscapular fossa | Internal rotation |
| Deltoid | Spine + acromion | Abduction (middle), flex/ext (anterior/posterior) |
| Biceps brachii (LH/SH) | Supraglenoid tubercle / coracoid | Flexion, supination |
| Triceps brachii (LH) | Infraglenoid tubercle | Extension |
| Coracobrachialis | Coracoid tip | Flexion/adduction of arm |
Neurovascular Structures
Suprascapular nerve (C5–C6, from superior trunk of brachial plexus):
- Passes under the transverse scapular ligament through the suprascapular notch → supplies supraspinatus
- Winds around the spinoglenoid notch under the spinoglenoid ligament → supplies infraspinatus
- The suprascapular artery passes over the transverse ligament (mnemonic: "navy over the bridge, army under")
Dorsal scapular nerve (C5): supplies levator scapulae and rhomboids, runs ~1–2 cm medial to the vertebral border
Long thoracic nerve (C5–C7): runs on the anterior surface of serratus anterior along the lateral thoracic wall — damage causes winged scapula (serratus anterior paralysis → medial border and inferior angle lift away from thorax)
Spinal accessory nerve (CN XI): innervates trapezius; travels with the superficial branch of the transverse cervical artery
Scapulothoracic Motion
The scapula moves through three axes (YXZ Euler sequence):
- Internal/external rotation (Y-axis) = protraction / retraction
- Upward/downward tilt (Z-axis, parallel to spine) = elevation / depression
- Lateral/medial rotation (X-axis) = glenoid moving superolaterally or inferomedially
With full arm elevation the scapula elevates 10–12 cm and the inferior angle translates laterally 10–15 cm with ~45–60° of tilt. The classic scapulohumeral rhythm approximates 2:1 glenohumeral to scapulothoracic motion.
Clinical Relevance
| Condition | Mechanism | Sign |
|---|---|---|
| Winged scapula | Long thoracic nerve injury (e.g., mastectomy, chest wall surgery, viral neuritis) → serratus anterior paralysis | Medial border + inferior angle protrude posteriorly, worsened on arm elevation or pushing against wall |
| Suprascapular nerve entrapment | Compression at suprascapular notch (e.g., ligament ossification in 10%) or spinoglenoid notch (ganglion cyst) | Shoulder pain + weakness of supraspinatus/infraspinatus; posterior shoulder atrophy |
| Os acromiale | Non-fusion of acromial ossification centers by age 25 | 7–15% incidence; can cause impingement or chronic shoulder pain |
| Scapular fractures | High-energy trauma (direct blow, humeral head impact, dislocation, violent muscular contracture) — associated with thoracic injuries; body and upper scapula most frequently affected |
Sources: Gray's Anatomy for Students; Imaging Anatomy Vol. 3 (Bones, Joints, Vessels & Nerves); Rockwood & Green's Fractures in Adults, 10th ed.; Miller's Review of Orthopaedics, 9th ed.
Innate immunity
Reading File
Finding Sources
Searching Images
innate immunity overview diagram pattern recognition receptors
Reading File
Reading File
Loading Image
Reading File
Finding Sources
Loading Image
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Loading Image
Finding Sources
Finding Sources
Finding Sources
Reading File
I now have comprehensive material from multiple authoritative sources. Let me compile the full answer.
Innate Immunity
Innate immunity is the first line of defense against infection, responding within minutes to hours of pathogen encounter. Unlike adaptive immunity, it does not require prior exposure and does not generate immunological memory. Its receptors recognize conserved structures shared among broad classes of pathogens rather than specific antigenic epitopes.
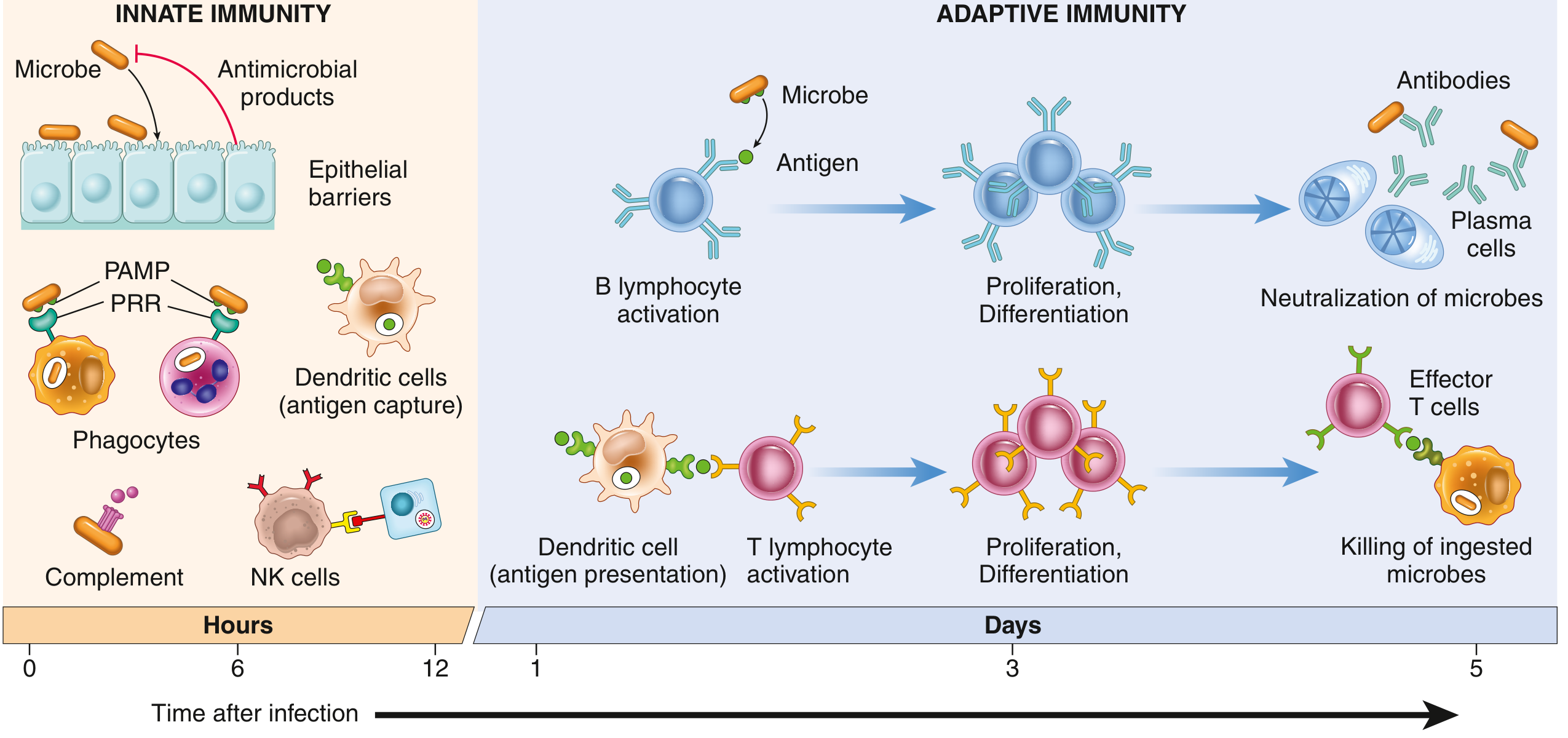
Lines of Defense
Human hosts have four interlocking tiers of protection:
- Natural barriers — skin, mucus, ciliated epithelium, gastric acid, bile
- Normal flora competition — resident microbiota inhibit pathogen colonization
- Innate immune responses — rapid, antigen-nonspecific (this topic)
- Adaptive immune responses — antigen-specific; reinforces innate responses; has memory
1. Physical and Chemical Barriers
Epithelial barriers (skin, GI, respiratory mucosa) act as mechanical obstacles. They also produce:
- Defensins — antimicrobial peptides that disrupt microbial membranes
- Antimicrobial chemicals — lysozyme, lactoferrin, secretory IgA (the latter is technically adaptive but constitutively present)
- Lung surfactant — coats and limits inhaled microbes
Lymphocytes residing within epithelia (intraepithelial lymphocytes) provide an additional rapid response at these surfaces.
2. Pattern Recognition Receptors (PRRs)
The central molecular logic of innate immunity: germline-encoded receptors recognizing conserved microbial structures.
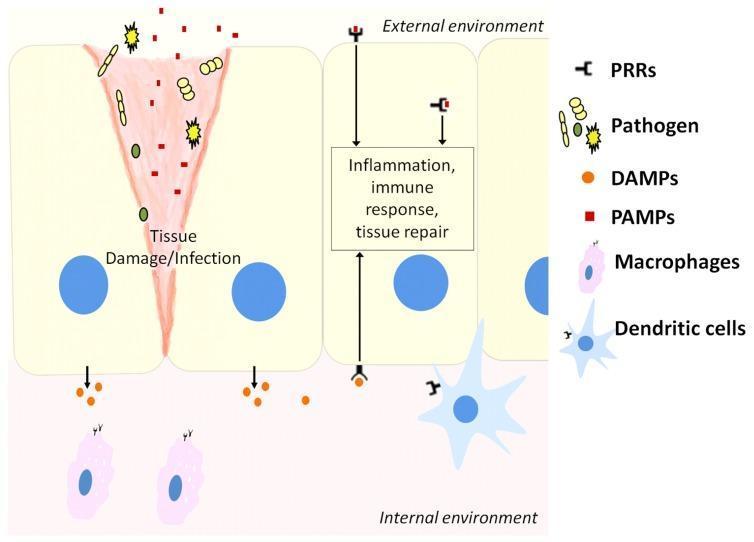
Ligands Recognized
| Signal | Source | Examples |
|---|---|---|
| PAMPs (Pathogen-associated molecular patterns) | Microbes | LPS, peptidoglycan, flagellin, viral RNA/DNA |
| DAMPs (Damage-associated molecular patterns) | Necrotic/injured host cells | Uric acid, ATP, HMGB1, cytosolic DNA, K⁺ efflux |
Innate immunity uses ~100 different receptors to recognize several thousand molecular patterns.
Classes of PRRs
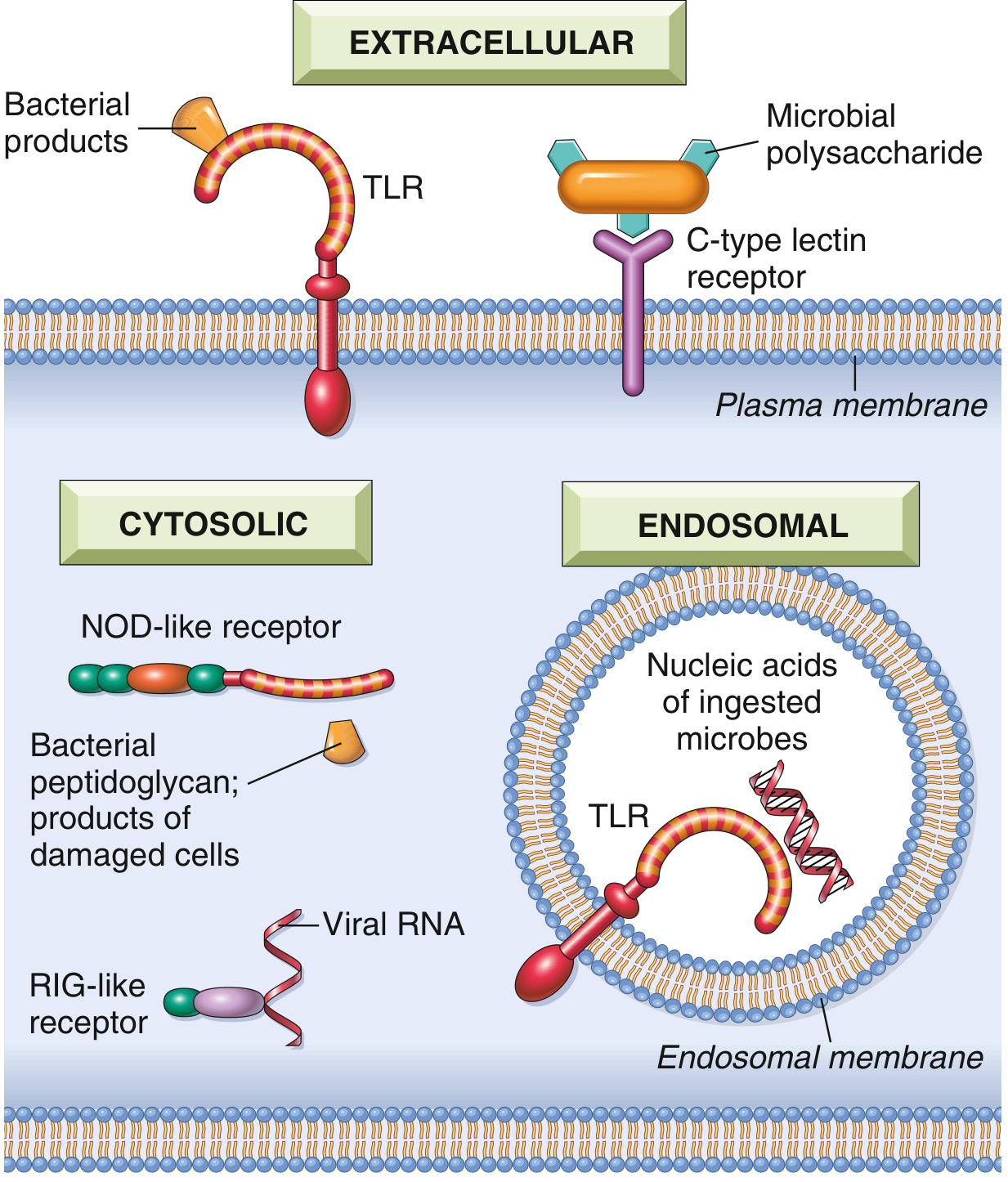
| Receptor Class | Location | Ligand | Outcome |
|---|---|---|---|
| Toll-like receptors (TLRs) | Plasma membrane (TLR1,2,4,5,6) + endosomes (TLR3,7,8,9) | LPS (TLR4), flagellin (TLR5), viral dsRNA (TLR3), CpG DNA (TLR9) | NF-κB activation → cytokines, interferons, costimulators |
| NOD-like receptors (NLRs) | Cytosol | Bacterial peptidoglycan fragments, DAMPs (uric acid, ATP, K⁺ loss) | Inflammation, inflammasome assembly |
| RIG-like receptors (RLRs) | Cytosol | Viral dsRNA (RIG-I, MDA5) | Type I IFN production; antiviral defense |
| C-type lectin receptors | Plasma membrane (macrophages, DCs) | Microbial polysaccharides (bacteria, fungi) | Phagocytosis, inflammatory cytokines |
| Cytosolic DNA sensors (cGAS-STING) | Cytosol | Foreign or aberrant host cytosolic DNA | Type I IFN production; interferonopathies if dysregulated |
| G protein-coupled receptors | Neutrophils, macrophages | N-formylmethionyl peptides (bacterial proteins) | Chemotaxis, activation |
3. Cellular Components
Phagocytes
Neutrophils — first recruited to sites of infection; rapidly phagocytose and kill microbes via:
- Reactive oxygen species (respiratory burst; NADPH oxidase)
- Lysosomal enzymes (myeloperoxidase, elastase)
- Neutrophil extracellular traps (NETs)
Macrophages — tissue-resident (Kupffer cells in liver, microglia in brain, alveolar macrophages in lungs) or monocyte-derived; perform:
- Phagocytosis and intracellular killing
- Cytokine production (TNF-α, IL-1, IL-6, IL-12)
- Antigen presentation to initiate adaptive immunity
Dendritic Cells (DCs)
- Sentinel cells in epithelia, lymphoid organs, and most tissues
- Richly equipped with PRRs; respond to microbes and cell damage
- Key bridge between innate and adaptive immunity: capture antigens → migrate to lymph nodes → present to naïve T cells → initiate adaptive response
- Produce cytokines directing T helper cell differentiation (IL-12 → Th1; IL-4 → Th2; IL-23 → Th17)
Natural Killer (NK) Cells
- Innate lymphocytes that kill virus-infected cells and tumor cells without prior sensitization
- Activated by cytokines (IL-12, IL-15, IL-18) from macrophages and DCs
- Missing-self hypothesis: NK cells constitutively express inhibitory receptors for self MHC class I; cells that downregulate MHC I (e.g., virally infected or malignant cells) lose this inhibitory signal and are killed
- Also carry activating receptors recognizing stress-induced ligands on infected/transformed cells
- Produce IFN-γ, which activates macrophages to kill phagocytosed microbes
Innate Lymphoid Cells (ILCs)
Tissue-resident lymphocytes lacking T-cell receptors; activated by cytokines rather than antigen:
| Group | Cytokines Produced | Functional Parallel |
|---|---|---|
| ILC1 | IFN-γ, TNF | Th1 |
| ILC2 | IL-4, IL-5, IL-13 | Th2 |
| ILC3 | IL-17, IL-22 | Th17 |
Mast Cells
- Tissue-resident; rich in preformed granules
- PRRs recognize PAMPs → degranulation → histamine, prostaglandins, leukotrienes
- Important at mucosal surfaces (respiratory, GI tracts)
Innate T Cells
γδ T cells, NKT cells, MAIT cells — bridge innate and adaptive systems; respond to conserved microbial lipids, phosphoantimetabolites, and vitamin B derivatives (not classical peptide antigens).
4. The Inflammasome
A cytosolic multiprotein complex assembled by certain NLRs (especially NLRP3):
- NLRs sense danger signals (urate crystals, ATP, K⁺ efflux, microbial products)
- Inflammasome recruits caspase-1
- Caspase-1 cleaves pro-IL-1β and pro-IL-18 → active IL-1β and IL-18 (potent pro-inflammatory cytokines)
- Caspase-1 also cleaves gasdermin D → membrane pores → pyroptosis (inflammatory cell death)
- Non-canonical pathway: cytosolic LPS activates caspase-4/5 (human) or caspase-11 (mouse) → gasdermin D pores → NLRP3 activation
Clinical relevance:
- Gout/pseudogout — urate/calcium pyrophosphate crystals activate NLRP3
- Gain-of-function NLR mutations → autoinflammatory syndromes (respond to IL-1 antagonists, e.g., anakinra)
- Dysregulation implicated in atherosclerosis, NAFLD, T2DM, Alzheimer disease
5. The Complement System
Over 30 plasma proteins (mostly synthesized by the liver), circulating as inactive zymogens, activated by three pathways:
| Pathway | Trigger | Key Initiator |
|---|---|---|
| Alternative | Pathogen surface alone (spontaneous C3 hydrolysis) | C3b deposition |
| Lectin | Mannose-binding lectin (MBL) or ficolins binding carbohydrates on pathogens | MASP-1/2 |
| Classical | Antibody–antigen complexes (bridges innate/adaptive) | C1q |
All three converge on C3 convertase → cleaves C3 → three effector arms:
- Opsonization — C3b coats microbes → phagocytosis via complement receptors (CR1, CR3)
- Inflammation — C3a and C5a (anaphylatoxins) → mast cell degranulation, neutrophil recruitment, vascular permeability
- Membrane attack complex (MAC) — C5b-9 → pore formation → direct lysis (especially encapsulated bacteria, Neisseria spp.)
Regulation — factor H, factor I, DAF (CD55), CD59 protect host cells from complement attack.
6. Soluble Mediators
| Mediator | Source | Function |
|---|---|---|
| TNF-α | Macrophages, DCs | Fever, acute-phase response, leukocyte recruitment, septic shock |
| IL-1β | Macrophages (inflammasome) | Fever, acute-phase proteins, T cell activation |
| IL-6 | Macrophages, endothelium | Fever, hepatic acute-phase response |
| IL-12 | DCs, macrophages | NK cell activation; induces IFN-γ; bridges to Th1 adaptive response |
| IFN-α/β (Type I IFNs) | Infected cells (via RLRs, cGAS-STING, endosomal TLRs) | Inhibit viral replication; upregulate MHC class I |
| IFN-γ (Type II IFN) | NK cells, ILC1 | Macrophage activation; Th1 polarization |
| Chemokines (CXCL8/IL-8) | Macrophages, endothelium | Neutrophil chemotaxis |
| Mannose-binding lectin | Liver | Opsonin; activates lectin complement pathway |
| C-reactive protein (CRP) | Liver (acute phase) | Binds phosphocholine on bacteria/dead cells → classical complement |
7. Innate Immunity and Inflammation
Innate immune recognition triggers the vascular and cellular response of acute inflammation:
- Vasodilation, increased permeability (histamine, prostaglandins, C3a/C5a)
- Leukocyte margination and emigration (selectins, integrins, chemokines)
- Recruited neutrophils and macrophages destroy pathogens and clear debris
8. Linking Innate to Adaptive Immunity
The innate system generates three essential signals for adaptive immune activation:
- Antigen — DCs phagocytose microbes, process proteins, and present peptides on MHC to T cells
- Costimulators — TLR activation upregulates B7 molecules (CD80/CD86) on DCs → engage CD28 on T cells (signal 2)
- Cytokines — IL-12, IL-4, IL-6, TGF-β drive T helper subset polarization (Th1/Th2/Th17/Treg)
Without innate immune activation (signals 2 and 3), antigen presentation alone leads to T cell anergy rather than activation — the basis for peripheral tolerance to self-antigens.
Distinguishing Features vs. Adaptive Immunity
| Feature | Innate | Adaptive |
|---|---|---|
| Speed | Minutes–hours | Days–weeks |
| Receptor diversity | ~100 PRRs (germline encoded) | 10⁷–10⁹ specificities (somatic rearrangement) |
| Memory | None (classically) | Yes (memory B and T cells) |
| Specificity | Conserved microbial patterns | Individual epitopes |
| Major cells | Phagocytes, NK cells, DCs, ILCs | T lymphocytes, B lymphocytes |
| Effector proteins | Complement, defensins, cytokines | Antibodies, cytokines |
Note on trained immunity: Recent evidence shows innate cells (especially monocytes/macrophages, NK cells) can undergo epigenetic reprogramming after initial activation, leading to enhanced responses upon re-exposure — a phenomenon called trained immunity. This challenges the classical dogma that innate immunity has no memory.
Sources: Cellular and Molecular Immunology (Abbas & Lichtman); Robbins & Kumar Basic Pathology; Robbins, Cotran & Kumar Pathologic Basis of Disease; Janeway's Immunobiology, 10e; Medical Microbiology, 9e
Mechanism of innate immunity
Finding Sources
Searching Images
innate immunity signaling mechanism TLR NF-kB pathway diagram
Your credits are exhausted. Your daily credits reset tomorrow, or you can purchase a top-up for more.
This is a shared conversation. Sign in to Orris to start your own chat.