What is bainbridge reflex
Reading File
Finding Sources
Searching PubMed
Bainbridge reflex heart rate atrial stretch
Reading File
Reading File
Reading File
Reading File
Here is a detailed, textbook-based explanation of the Bainbridge reflex:
Bainbridge Reflex
Definition
The Bainbridge reflex is the tachycardia (increase in heart rate) produced by an increase in venous return and atrial pressure (volume loading). It was named after the British physiologist Francis Arthur Bainbridge who described it in 1915.
Stimulus
- Increased blood volume or venous return causes distension of the right atrium and great veins
- This stretches low-pressure B-type stretch receptors (mechanoreceptors) in the atrial walls
Reflex Arc
| Component | Details |
|---|---|
| Receptor | Low-pressure atrial stretch receptors (B fibers) in the atrial walls |
| Afferent limb | Signals travel via the vagus nerve to the nucleus tractus solitarius (NTS) in the medulla |
| Efferent limb | Response is carried back to the SA node via both vagal (parasympathetic) and sympathetic pathways |
| Effector | SA node - increases heart rate; minor effects on contractility |
Physiological Response
- Heart rate increases up to 75% (40-60% via the Bainbridge reflex + 10-15% by direct stretch of the SA node)
- Efferent signals also increase strength of cardiac contraction modestly
- The reflex saturates at high baseline heart rates - the increase is greatest when the resting heart rate is slow
Purpose / Significance
The Bainbridge reflex prevents blood from pooling in the veins, atria, and pulmonary circulation. When venous return is high, the heart must pump faster to "keep up" and clear the incoming blood. It acts as a complementary mechanism alongside:
- Frank-Starling law (more filling = more force)
- Baroreceptor reflex (arterial pressure regulation)
Bainbridge Reflex vs. Baroreceptor Reflex - Competition
This is a key physiological concept. Both reflexes respond to volume changes, but in opposite directions for the same stimulus:
| Situation | Bainbridge Reflex | Baroreceptor Reflex | Net Effect on Heart Rate |
|---|---|---|---|
| Volume loading (high blood volume) | Increases HR | Decreases HR (high BP activates baroreceptors) | Bainbridge dominates → HR rises |
| Volume depletion (low blood volume) | Minimal effect | Increases HR (low BP reduces baroreceptor firing) | Baroreceptor reflex dominates → HR rises |
| Normal volume | Neither strongly activated | Baseline | HR is at its minimum |
This explains why both volume loading and volume depletion cause tachycardia, but by different mechanisms. Heart rate is at its lowest when circulating volume is normal.
Key Points to Remember
- The Bainbridge reflex is a low-pressure receptor reflex (not arterial baroreceptors)
- The efferent limb involves both sympathetic and parasympathetic pathways to the SA node
- It primarily affects heart rate, not significantly contractility or stroke volume
- The reflex saturates - effective mainly when baseline HR is low
- Works in concert with the Frank-Starling mechanism and baroreceptor reflex to regulate cardiac output
Sources: Guyton and Hall Textbook of Medical Physiology; Medical Physiology (Boron & Boulpaep); Costanzo Physiology, 7th Edition
Chemiosmotic hypothesis
Finding Sources
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Chemiosmotic Hypothesis (Mitchell Hypothesis)
Proposed by Peter Mitchell in 1961 - Nobel Prize in Chemistry, 1978. It explains how the free energy generated by the electron transport chain (ETC) is used to produce ATP from ADP + Pi during oxidative phosphorylation.
Core Concept
The hypothesis states that the energy for ATP synthesis is provided by an electrochemical (proton) gradient across the inner mitochondrial membrane - NOT by a direct chemical intermediate as previously believed.
Step-by-Step Mechanism
Step 1 - Electron Transport and Proton Pumping
Electrons donated by NADH and FADH2 (from the citric acid cycle) pass through the ETC complexes embedded in the inner mitochondrial membrane:
| Complex | Name | Protons Pumped (per NADH) |
|---|---|---|
| Complex I | NADH dehydrogenase | 4 H+ |
| Complex II | Succinate dehydrogenase | 0 H+ (no pumping) |
| Complex III | Cytochrome bc1 complex | 4 H+ |
| Complex IV | Cytochrome c oxidase | 2 H+ |
| Total | Per NADH oxidized | 10 H+ pumped |
- At the end of the chain, O2 is the final electron acceptor, reduced to H2O
- Electrons from FADH2 enter at Complex II, yielding only 6 H+ (bypassing Complex I)
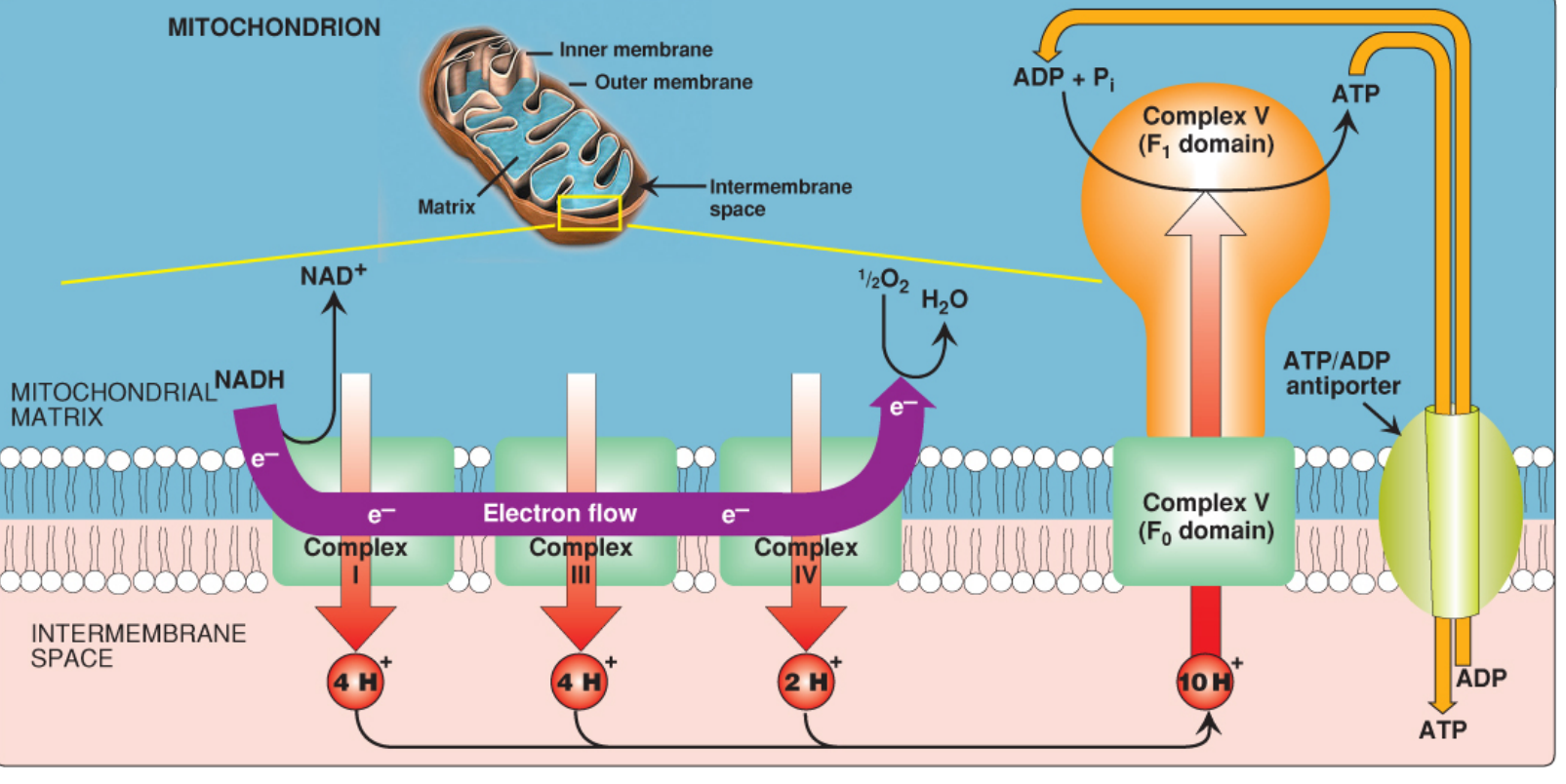
Step 2 - Formation of the Proton Motive Force (PMF)
The continuous pumping of H+ from the matrix → intermembrane space (IMS) creates two gradients:
- Chemical gradient (pH gradient, ΔpH) - IMS is more acidic (~0.75 pH units lower) than the matrix
- Electrical gradient (membrane potential, Δψ) - IMS is positively charged relative to the matrix
Together, these form the Proton Motive Force (PMF) - the combined electrochemical driving force that pushes protons back into the matrix.
The inner mitochondrial membrane is impermeable to protons, so they cannot simply diffuse back - they can only return through a specific channel (ATP synthase).
Step 3 - ATP Synthesis via ATP Synthase (Complex V / F0F1-ATPase)
H+ ions flow back down the electrochemical gradient through ATP synthase, and this flow drives ATP synthesis.
ATP synthase has two functional domains:
| Domain | Location | Function |
|---|---|---|
| F0 | Spans the inner mitochondrial membrane | Contains the H+ channel (c-ring subunits + subunit a) |
| F1 | Protrudes into the mitochondrial matrix | Contains the catalytic sites (3 αβ subunit pairs); synthesizes ATP |
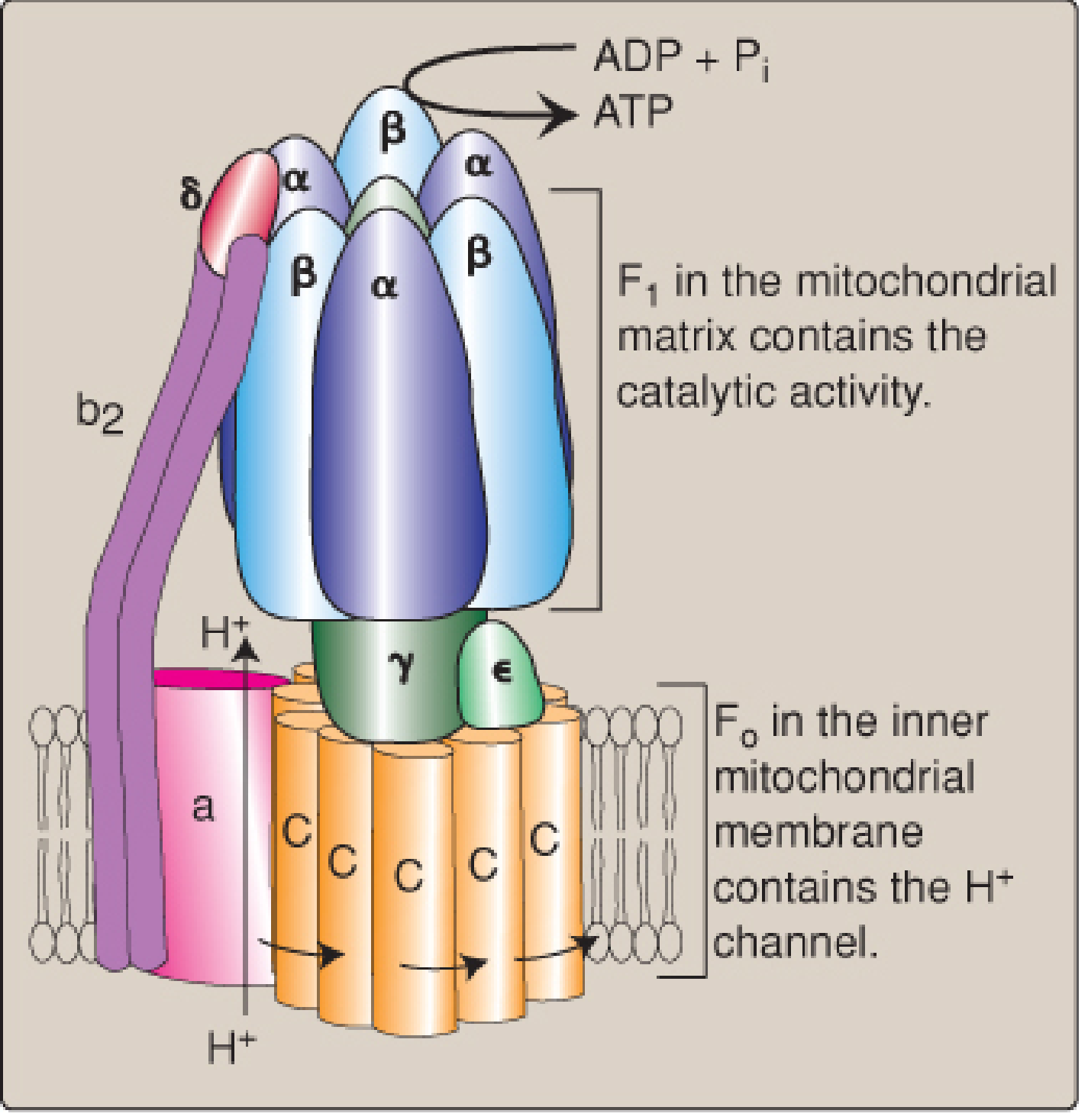
Mechanism of rotation (Binding Change Mechanism):
- H+ enters from the IMS into a channel in the F0 domain
- Each H+ protonates a glutamyl carboxyl group on a c-subunit, causing the c-ring to rotate
- Rotation of the c-ring turns the γ-shaft (stalk), which is connected to the F1 domain
- As the γ-shaft rotates, it changes the conformation of the three β-subunits in F1 sequentially through three states: Open (O), Loose (L), Tight (T)
- In the Tight conformation, ADP + Pi are spontaneously phosphorylated to ATP
- Rotation to the Open conformation causes ATP release
- One complete rotation of the c-ring = 3 ATP synthesized
- In vertebrates, the c-ring has 8 subunits, so 8 H+ per full rotation = 3 ATP → ~2.7 H+ per ATP
Summary of Coupling
NADH/FADH2 → ETC → H+ pumped out → PMF created → H+ flows through F0F1 → ATP synthesized
(oxidation) (gradient) (phosphorylation)
This is why it is called oxidative phosphorylation - oxidation of NADH/FADH2 is directly coupled to phosphorylation of ADP via the H+ gradient as the common intermediate.
Inhibitors and Uncouplers
| Agent | Mechanism | Effect |
|---|---|---|
| Oligomycin | Blocks F0 H+ channel | Stops ATP synthesis AND electron transport (respiratory control) |
| Rotenone | Blocks Complex I | Inhibits NADH oxidation |
| Cyanide / CO | Blocks Complex IV | Stops O2 reduction; halts entire chain |
| 2,4-Dinitrophenol (DNP) | Lipophilic H+ carrier (ionophore) | Shuttles H+ across membrane, dissipates gradient; energy released as heat |
| UCP1 (Thermogenin) | Uncoupling protein in brown fat | Natural uncoupler - allows H+ back-flux, generating heat (non-shivering thermogenesis) |
| Aspirin (toxic doses) | Uncoupler | Dissipates gradient, causes fever |
Uncouplers allow electron transport to continue at a rapid rate (consuming O2 and fuel) but without generating ATP - energy is lost as heat.
Key Points
- The H+ gradient is the common intermediate that links oxidation to phosphorylation
- The inner mitochondrial membrane must be intact and impermeable for the hypothesis to work
- Coupling: electron transport and ATP synthesis are tightly linked through the H+ gradient; increasing ADP availability increases H+ flow, which accelerates electron transport
- The same principle applies in chloroplasts (light-driven proton pumping across the thylakoid membrane drives ATP synthesis in photosynthesis)
Sources: Biochemistry 8th ed, Lippincott Illustrated Reviews; Basic Medical Biochemistry - A Clinical Approach, 6th ed; Medical Physiology (Boron & Boulpaep)
This is a shared conversation. Sign in to Orris to start your own chat.