Explain each and everything about Electron transport chain
Reading File
Finding Sources
Searching the Web
electron transport chain complexes mechanism ATP synthesis
Reading File
Reading File
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Loading Image
I now have all the information needed from authoritative textbooks plus excellent diagrams. Let me compose the comprehensive answer.
The Electron Transport Chain (ETC) - Complete Guide
Sources: Biochemistry, 8th ed. Lippincott Illustrated Reviews; Basic Medical Biochemistry - A Clinical Approach, 6th ed.
1. Overview and Location
The ETC (also called the respiratory chain) is the final common pathway by which electrons derived from different fuels flow to oxygen, reducing it to water. It is located in the inner mitochondrial membrane (except cytochrome c, which is in the intermembrane space). The coupling of electron transport with ATP synthesis is called oxidative phosphorylation (OXPHOS).
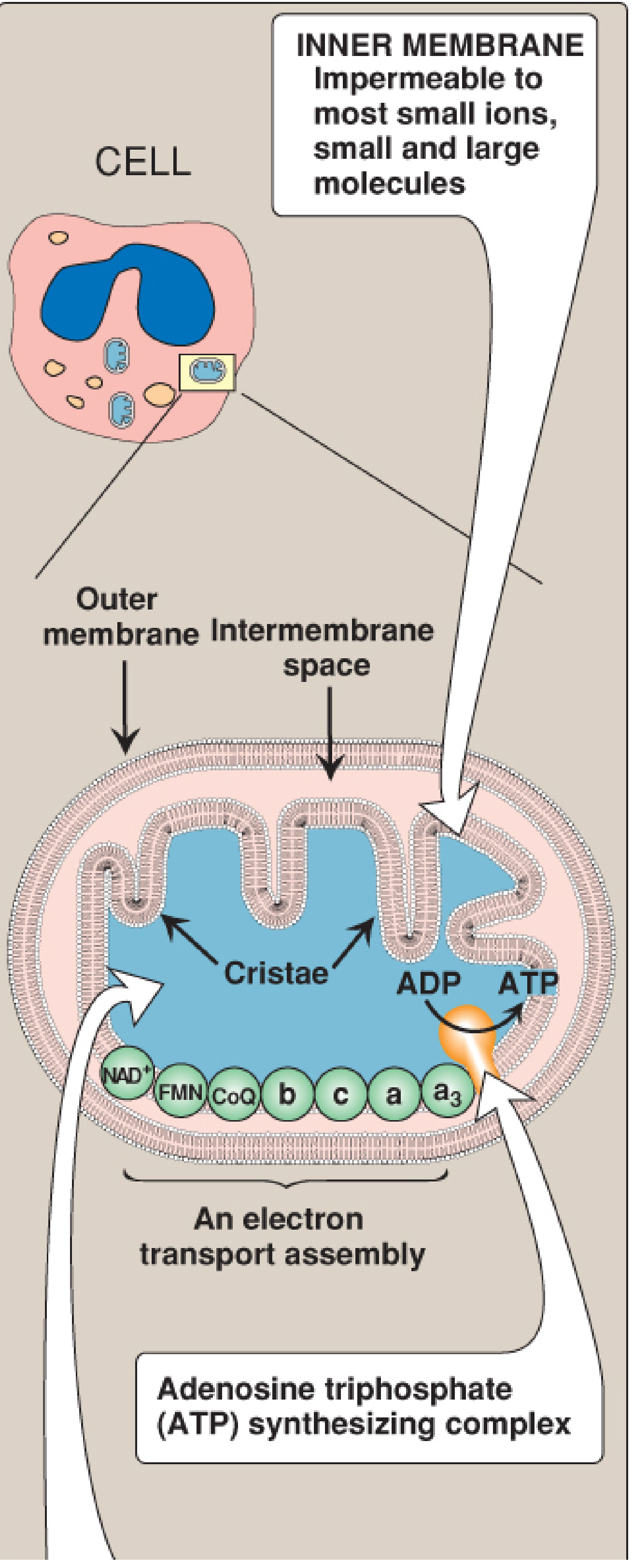
Figure: Mitochondrial structure - the ETC and ATP synthase sit on the inner membrane, which is impermeable to most ions and small molecules. The cristae greatly increase its surface area. - Biochemistry, 8th ed. Lippincott, p. 226
2. The Mitochondrial Membranes
| Membrane | Permeability | Function |
|---|---|---|
| Outer membrane | Freely permeable (protein porin forms channels) | Allows ions and small molecules to pass |
| Intermembrane space | Similar composition to cytosol | H+ accumulates here during ETC |
| Inner membrane | Impermeable to most ions, ATP, ADP, pyruvate | Houses ETC complexes; requires transport proteins |
| Matrix | Gel-like interior | Site of TCA cycle, fatty acid oxidation, mtDNA |
The inner mitochondrial membrane is unusually rich in proteins - over half are directly involved in oxidative phosphorylation. The convolutions called cristae greatly increase its surface area. - Biochemistry, 8th ed. Lippincott, p. 226
3. Electron Donors: NADH and FADH2
Energy-rich molecules like glucose are metabolized by oxidation reactions yielding CO2 and water. The metabolic intermediates donate electrons to two key coenzymes:
- NAD+ is reduced to NADH (carries a hydride ion, H-)
- FAD is reduced to FADH2 (carries two hydrogen atoms)
These reduced coenzymes then donate electron pairs to the ETC. The electrons lose free energy as they pass down the chain, and this energy drives proton pumping and ultimately ATP synthesis. - Biochemistry, 8th ed. Lippincott, p. 224
4. The Four Complexes + Mobile Carriers
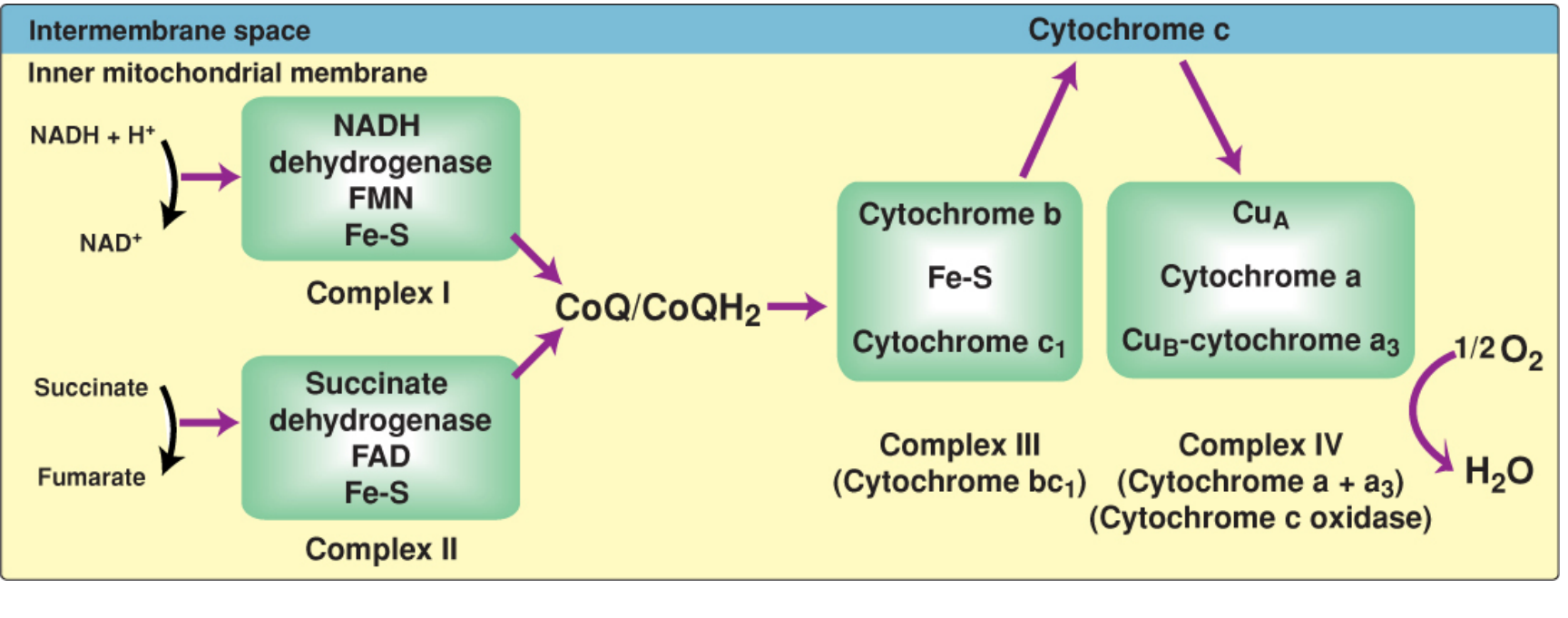
Figure: Complete electron transport chain. Complex I (NADH dehydrogenase) and Complex II (succinate dehydrogenase) both feed electrons to CoQ. Complex III passes them to cytochrome c, and Complex IV reduces O2 to H2O. - Biochemistry, 8th ed. Lippincott, p. 232
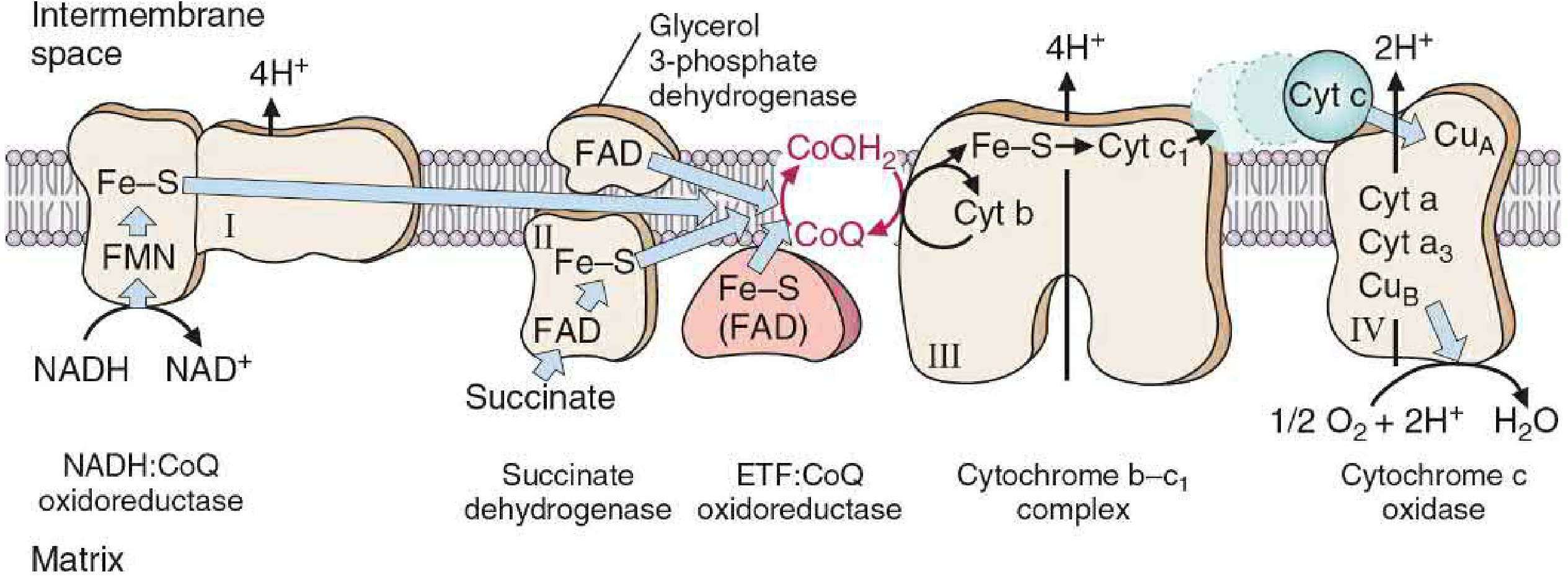
Figure: Oxidation-reduction components of the ETC. Note that only Complexes I, III, and IV pump protons. Complex II, glycerol-3-phosphate DH, and ETF:CoQ oxidoreductase all donate to CoQ but do NOT pump protons. - Basic Medical Biochemistry, 6th ed., p. 869
Complex I - NADH Dehydrogenase (NADH:CoQ Oxidoreductase)
- Also called: NADH dehydrogenase
- Size: ~45 subunit complex - one of the largest known protein complexes
- Components: FMN (flavin mononucleotide), multiple Fe-S centers, CoQ binding sites
- Reaction: NADH + H+ + CoQ → NAD+ + CoQH2
- Electron path: NADH → FMN → Fe-S centers → CoQ
- Protons pumped: 4 H+ per electron pair, from matrix to intermembrane space
- Inhibitors: Rotenone (pesticide), Amytal (amobarbital), MPTP (neurotoxin)
FMN accepts two electrons from NADH but can pass only single electrons to the Fe-S centers. Fe-S centers can delocalize single electrons into large orbitals, allowing electron transfer to and from CoQ. - Basic Medical Biochemistry, 6th ed., p. 872
Complex II - Succinate Dehydrogenase
- Also called: Succinate:CoQ oxidoreductase
- Unique feature: The only ETC complex that is also a TCA cycle enzyme
- Components: FAD, Fe-S protein
- Reaction: Succinate + FAD → Fumarate + FADH2 → CoQ
- Electron path: Succinate → FAD → Fe-S → CoQ
- Protons pumped: 0 H+ (does NOT pump protons - no energy is lost in this transfer; it does not span the membrane with a pumping mechanism)
- Also accepts from: Glycerol-3-phosphate dehydrogenase, ETF:CoQ oxidoreductase (from fatty acid beta-oxidation via electron-transferring flavoprotein)
- Inhibitors: Malonate (competitive inhibitor of succinate binding)
Because no energy is lost in this process, no H+ are pumped at Complex II. - Biochemistry, 8th ed. Lippincott, p. 229
Mobile Carrier: Coenzyme Q (CoQ / Ubiquinone)
- A lipid-soluble quinone with a long hydrophobic isoprenoid tail (derived from cholesterol synthesis intermediate)
- Freely diffusible in the lipid bilayer - acts as a mobile electron shuttle
- Collects electrons from Complex I, Complex II, and other flavoprotein dehydrogenases
- Transfers electrons to Complex III
- Exists in three forms: CoQ (oxidized), CoQ semiquinone (CoQ•-, one electron), CoQH2 (reduced, hydroquinone)
- The Q cycle at Complex III involves these three redox states
CoQ is a mobile electron carrier and can accept electrons from NADH dehydrogenase, succinate dehydrogenase, glycerol-3-phosphate dehydrogenase, and acyl CoA dehydrogenases. - Biochemistry, 8th ed. Lippincott, p. 230
Complex III - Cytochrome bc1 Complex (CoQ:Cytochrome c Oxidoreductase)
- Components: Cytochrome b, Fe-S center (Rieske protein), cytochrome c1
- Reaction: CoQH2 + 2 cytochrome c (Fe3+) → CoQ + 2 cytochrome c (Fe2+) + 4 H+
- Mechanism: The Q cycle - CoQH2 releases 2 electrons; one goes through Fe-S → cyt c1 → cyt c; the other reduces cytochrome b and regenerates CoQ semiquinone
- Protons pumped: 4 H+ per electron pair
- Inhibitors: Antimycin A (binds cytochrome b)
Mobile Carrier: Cytochrome c
- A small, water-soluble heme protein in the intermembrane space
- Loosely associated with the outer face of the inner membrane
- Carries single electrons (Fe2+ ↔ Fe3+)
- Shuttles electrons from Complex III to Complex IV
- Unlike hemoglobin, its heme iron is reversibly oxidized/reduced as part of its normal function
- Also plays a role in apoptosis when released from the intermembrane space into the cytosol
Complex IV - Cytochrome c Oxidase (Cytochrome a + a3)
- Also called: Cytochrome c oxidase
- Components: CuA, cytochrome a, cytochrome a3, CuB
- Reaction: 4 cytochrome c (Fe2+) + O2 + 4 H+ → 4 cytochrome c (Fe3+) + 2 H2O
- Electron path: Cyt c → CuA → cyt a → cyt a3 (with CuB) → O2
- Protons pumped: 2 H+ per electron pair
- Special feature: The ONLY complex where heme iron has an available coordination site that can react directly with O2, making it the terminal oxidase
- Requires 4 electrons to fully reduce one O2 to two H2O (prevents toxic partial reduction to superoxide)
- Inhibitors: Cyanide (CN-), carbon monoxide (CO), hydrogen sulfide (H2S), sodium azide (NaN3) - all bind cytochrome a3 and block O2 reduction
- Proton movement mechanism: Involves direct transport through a series of bound water molecules or amino acid side chains - described as a "proton wire"
Cytochrome c oxidase is the only electron carrier in which heme iron has an available coordination site that can react directly with O2. - Biochemistry, 8th ed. Lippincott, p. 233
5. Summary: Proton Pumping at Each Complex
| Complex | Protons Pumped (H+/pair) | Inhibitors |
|---|---|---|
| Complex I | 4 | Rotenone, Amytal, MPTP |
| Complex II | 0 | Malonate |
| Complex III | 4 | Antimycin A |
| Complex IV | 2 | CN-, CO, H2S, NaN3 |
| Total (NADH) | 10 | |
| Total (FADH2) | 6 (bypasses Complex I) |
6. Free Energy and Reduction Potentials
Electrons flow spontaneously from lower to higher reduction potential (E0). The more negative the E0, the stronger the electron donor:
| Redox Pair | E0 (V) |
|---|---|
| NADH/NAD+ | -0.32 V (strong donor) |
| FADH2/FAD | -0.18 V |
| CoQH2/CoQ | +0.04 V |
| Cytochrome c (Fe2+/Fe3+) | +0.23 V |
| H2O/O2 | +0.82 V (strong acceptor) |
The relationship between free energy and reduction potential:
ΔG° = -nFΔE0
where n = number of electrons, F = Faraday constant (23.1 kcal/volt·mol)
- Transport of 2 electrons from NADH to O2 releases 52.6 kcal of free energy
- Phosphorylation of ADP to ATP costs +7.3 kcal/mol
- The chain runs irreversibly from high-energy electrons (NADH) to the low-energy final acceptor (O2)
7. Chemiosmotic Hypothesis (Mitchell Hypothesis) and ATP Synthesis
Peter Mitchell proposed this in 1961 (Nobel Prize 1978). The key idea: electron transport does not directly synthesize ATP. Instead, it builds a proton gradient that drives ATP synthase.
The Proton Gradient
Proton pumping at Complexes I, III, and IV creates:
- Electrical gradient: More positive charges (H+) on the cytosolic/intermembrane side
- pH gradient: Intermembrane space is more acidic (lower pH) than the matrix
- Together these form the proton-motive force (PMF)
For each NADH oxidized, 10 H+ total are pumped across.
Complex V - ATP Synthase (F0F1-ATPase)
ATP synthase has two major components:
- F0 (membrane sector): Embedded in the inner membrane; consists of c-ring (rotor), a-subunit (proton channel), and b-subunits. The c-ring rotates as protons flow through the channel.
- F1 (catalytic headpiece): Protrudes into the matrix; contains 3 α and 3 β subunits forming the catalytic sites, plus γ (shaft/stalk) that rotates inside the αβ hexamer.
Binding Change Mechanism (Paul Boyer, Nobel Prize 1997):
The rotating γ-shaft alters the conformation of each β subunit sequentially through three states:
- Open (O): Low affinity - binds ADP + Pi
- Loose (L): Binds substrates, spontaneously forms ATP
- Tight (T): High affinity - ATP is trapped; rotation releases it
Each full rotation of the γ-shaft synthesizes 3 ATP. Each ATP release requires ~3 H+ (through the rotor). An additional H+ is used to transport Pi into the matrix, so effectively ~4 H+ per ATP.
ATP yield: ~2.5 ATP per NADH oxidized; ~1.5 ATP per FADH2 oxidized (modern accepted estimates). Only ~30% of available energy is captured as ATP - the rest generates heat.
8. Reactive Oxygen Species (ROS)
Electron leakage from the ETC (mostly at Complexes I and III) produces ROS:
| ROS | Formula |
|---|---|
| Superoxide | O2•- |
| Hydrogen peroxide | H2O2 |
| Hydroxyl radical | •OH |
ROS damages DNA, proteins, and causes lipid peroxidation. Cellular defenses include:
- Superoxide dismutase (SOD): O2•- → H2O2
- Catalase: H2O2 → H2O + O2
- Glutathione peroxidase: H2O2 + GSH → H2O + GSSG
9. Inhibitors and Uncouplers
Site-Specific Inhibitors
These block electron flow at specific complexes, causing a buildup of reduced carriers before the block and oxidized carriers after it. Since electron transport and ATP synthesis are tightly coupled, inhibiting the ETC also inhibits ATP synthesis.
| Inhibitor | Site of Action | Mechanism |
|---|---|---|
| Rotenone | Complex I | Blocks electron flow from Fe-S to CoQ |
| Amytal (amobarbital) | Complex I | Same as rotenone |
| MPTP | Complex I | Neurotoxin; causes Parkinson-like syndrome |
| Malonate | Complex II | Competitive inhibitor of succinate |
| Antimycin A | Complex III | Binds cytochrome b |
| Cyanide (CN-) | Complex IV | Binds cytochrome a3, blocks O2 reduction |
| Carbon monoxide (CO) | Complex IV | Same as cyanide |
| H2S | Complex IV | Same as cyanide |
| Sodium azide (NaN3) | Complex IV | Same as cyanide |
| Oligomycin | ATP synthase (F0) | Blocks proton channel of F0 |
Uncouplers
These dissociate electron transport from ATP synthesis by allowing H+ to re-enter the matrix without going through ATP synthase (they bypass Complex V). Electron transport continues (or even accelerates) but no ATP is made - energy is released as heat instead.
| Uncoupler | Details |
|---|---|
| 2,4-Dinitrophenol (DNP) | Lipid-soluble proton carrier; carries H+ across the inner membrane bypassing ATP synthase; used as a weight-loss drug historically - very dangerous |
| Thermogenin (UCP-1) | Physiological uncoupler in brown adipose tissue; allows H+ back-flow; produces heat for cold adaptation in neonates and hibernating animals |
DNP is shown in its reduced (DNPH) and oxidized (DNP-) forms as it cycles H+ across the membrane. - Biochemistry, 8th ed. Lippincott, p. 244
10. NADH Shuttle Systems
The inner mitochondrial membrane has no NADH transporter. Cytosolic NADH (from glycolysis) cannot directly enter. Two shuttle systems transfer its reducing equivalents:
Glycerol-3-Phosphate Shuttle
- Cytosolic NADH reduces DHAP → glycerol-3-phosphate
- Mitochondrial isozyme oxidizes glycerol-3-phosphate → FADH2
- Net result: 1.5 ATP per cytosolic NADH (FADH2 bypasses Complex I)
- Used primarily in brain and muscle
Malate-Aspartate Shuttle
- Cytosolic NADH reduces oxaloacetate → malate
- Malate enters mitochondrial matrix and is oxidized back to oxaloacetate → NADH
- Net result: 2.5 ATP per cytosolic NADH (NADH enters the full chain)
- Used primarily in heart, liver, and kidney
11. Energy Yield Calculation
| Substrate | ATP Yield (modern estimate) |
|---|---|
| 1 NADH | ~2.5 ATP |
| 1 FADH2 | ~1.5 ATP |
| 1 GTP (from TCA) | 1 ATP |
Per glucose molecule:
- Glycolysis: 2 ATP (substrate level) + 2 NADH (cytosolic, via shuttle: 3-5 ATP)
- Pyruvate dehydrogenase: 2 NADH → 5 ATP
- TCA cycle: 6 NADH → 15 ATP + 2 FADH2 → 3 ATP + 2 GTP → 2 ATP
- Total: ~30-32 ATP (modern estimate; older textbooks cite 36-38)
The overall free energy release:
- NADH oxidation: ~53 kcal/mol
- FADH2 oxidation: ~41 kcal/mol
- Only ~30% is captured as ATP; the rest is heat (making the ETC our major source of body heat)
12. Coupling and Control
The processes of electron transport and ATP synthesis are tightly coupled - one cannot occur without the other under normal conditions:
- When ADP is available → H+ flows through ATP synthase → proton gradient drops → ETC accelerates
- When ADP is scarce (cell is energy-replete) → H+ gradient builds up → "proton back pressure" slows ETC → O2 consumption falls
- This means ETC activity is controlled by the energy demands of the cell (respiratory control)
13. Mitochondrial DNA and Inherited Defects
- 13 of ~90 polypeptides required for OXPHOS are encoded by mitochondrial DNA (mtDNA)
- The remaining are encoded by nuclear DNA, synthesized in the cytosol, and imported
- mtDNA has a mutation rate ~10x greater than nuclear DNA
- Defects cause lactic acidosis (especially in muscles and CNS), because without ETC function, cells shift to anaerobic glycolysis
- High-energy-demand tissues are most vulnerable: brain, nerves, retina, skeletal and cardiac muscle, liver
Clinical Manifestations of OXPHOS Disorders
- Seizures
- Ophthalmoplegia (external eye muscle weakness)
- Muscle weakness / myopathy
- Cardiomyopathy
- Lactic acidosis
- Some medications (e.g., certain antiretrovirals) impair mitochondrial function
14. Clinical Relevance Summary
| Scenario | Mechanism |
|---|---|
| Cyanide poisoning | Inhibits Complex IV → no O2 reduction → ETC halts → no ATP → lactic acidosis, rapid death |
| CO poisoning | Same as cyanide |
| DNP toxicity | Uncoupling → hyperthermia, tachycardia, diaphoresis, metabolic acidosis |
| Iron deficiency | Reduces Fe-S centers and cytochromes → fatigue even before severe anemia |
| Metformin | Mild Complex I inhibition in the liver → reduces hepatic gluconeogenesis |
| MPTP/Parkinson's | Complex I inhibition in substantia nigra → neuronal death |
| Mitochondrial myopathies | mtDNA mutations → ragged red fibers on muscle biopsy (Gomori trichrome stain) |
Here is an excellent animation to visualize the full process:
Key takeaways:
- The ETC has 4 complexes + 2 mobile carriers (CoQ, cytochrome c) + ATP synthase (Complex V)
- Only Complexes I, III, and IV pump protons (not Complex II)
- Total of 10 H+ pumped per NADH; 6 per FADH2
- The proton gradient (proton-motive force) drives ATP synthase by chemiosmosis
- ~2.5 ATP per NADH; ~1.5 ATP per FADH2; ~30-32 ATP per glucose total
- Inhibitors block specific complexes; uncouplers bypass ATP synthase entirely
This is a shared conversation. Sign in to Orris to start your own chat.