anterior frontalle
Reading File
Finding Sources
Searching the Web
anterior fontanelle anatomy clinical significance neonatal
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Here is a complete overview of the Anterior Fontanelle (fonticulus anterior):
Anterior Fontanelle
Anatomy
The anterior fontanelle is the largest of the six fontanelles in the neonatal skull. It forms at the junction where four bones meet: the two frontal bones and the two parietal bones. The sutures that converge here are:
- Coronal suture (frontal meets parietal, bilaterally)
- Sagittal suture (between the two parietal bones)
- Metopic (frontal) suture (between the two frontal bones)
This gives it a characteristic diamond (rhombus) shape, which distinguishes it from the triangular posterior fontanelle.
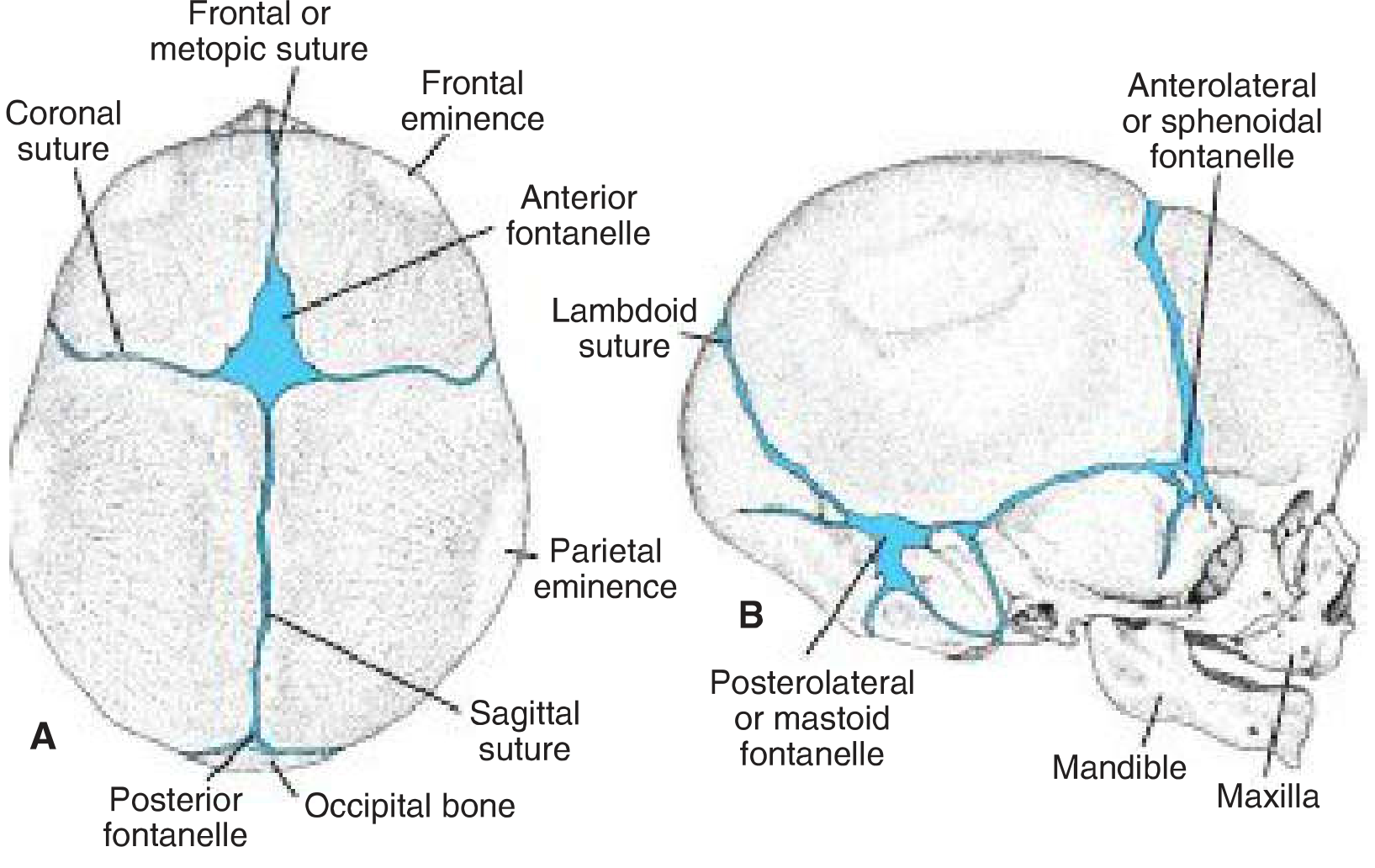
Skull of a newborn from above (A) and the right side (B). The anterior fontanelle is the large diamond-shaped gap at the intersection of the coronal, sagittal, and metopic sutures. - Langman's Medical Embryology
Size
- Range: 0.6 to 3.6 cm (mean ~2.1 cm in diameter)
- It is shaped like a cross (+), whereas the posterior fontanelle forms a Y-shape
- Significantly larger than all other fontanelles
Closure
| Fontanelle | Closure |
|---|---|
| Anterior | ~18 months of age (middle of the second year) |
| Posterior | 1-2 months after birth (some say by 8 weeks) |
| Sphenoidal (anterolateral) | ~3 months |
| Mastoid (posterolateral) | ~12-18 months |
Function
- Skull molding - allows flat bones to overlap during passage through the birth canal (a process called molding), protecting the brain during delivery
- Postnatal brain growth - sutures and fontanelles remain open to allow the cranial vault to expand as the brain grows (brain is ~66% of adult size by age 2)
- Clinical assessment window - palpation gives information about intracranial pressure and ossification progress
- Venipuncture access - in neonates and infants, the anterior fontanelle can be used to take blood samples from the dural sinuses (sagittal sinus) when peripheral access is limited - Color Atlas of Human Anatomy
Clinical Significance
Bulging anterior fontanelle (tense, full, pulsatile at rest):
- Raised intracranial pressure (ICP)
- Meningitis / encephalitis
- Hydrocephalus
- Intracranial hemorrhage
- Vitamin A toxicity
Sunken/depressed anterior fontanelle:
- Dehydration (most common cause in infants)
- Malnutrition
Large fontanelle or delayed closure (beyond 18-24 months):
- Down syndrome (trisomy 21)
- Achondroplasia
- Congenital hypothyroidism (cretinism)
- Rickets
- Elevated intracranial pressure
- Cleidocranial dysostosis
Premature closure (craniosynostosis):
- If the sagittal or coronal sutures fuse too early, the fontanelle closes prematurely and the skull cannot expand normally - compensatory growth may then occur through the anterior fontanelle (as in acrocephaly/oxycephaly) - Cummings Otolaryngology
Obstetric Relevance
During labor, identifying the shape and position of the anterior fontanelle helps determine fetal head position:
- Anterior fontanelle felt anteriorly in the pelvis = occiput posterior (OP) position
- Cruciform (+) shape of the anterior fontanelle vs. the Y-shape of the posterior fontanelle helps the clinician confirm presentation and station
In ENT/Rhinology (a different "anterior fontanelle"):
The term "anterior fontanelle" also appears in sinonasal anatomy - here it refers to a bony dehiscence (membranous area without bone) on the lateral nasal wall, located anterior to the articulation of the uncinate process with the inferior turbinate. Accessory maxillary ostia may form in this region. This is distinct from the neonatal skull fontanelle. - K.J. Lee's Essential Otolaryngology
development of somites
Finding Sources
Searching the Web
somite development embryology paraxial mesoderm segmentation
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Loading Image
Reading File
Reading File
Loading Image
Finding Sources
Here is a thorough, textbook-based account of somite development:
Development of Somites (Somitogenesis)
1. Origin - Paraxial Mesoderm
During gastrulation (week 3), cells migrating through the primitive streak form three germ layers. Those that migrate craniolaterally condense on each side of the notochord as paraxial mesoderm - a thick longitudinal column that runs the length of the embryo on either side of the developing neural tube.
- Paraxial mesoderm is continuous laterally with intermediate mesoderm, which thins into the lateral plate mesoderm
- The lateral plate splits into somatic (parietal) and splanchnic (visceral) layers by the intraembryonic coelom
2. Somitomeres - The Unsegmented Precursors
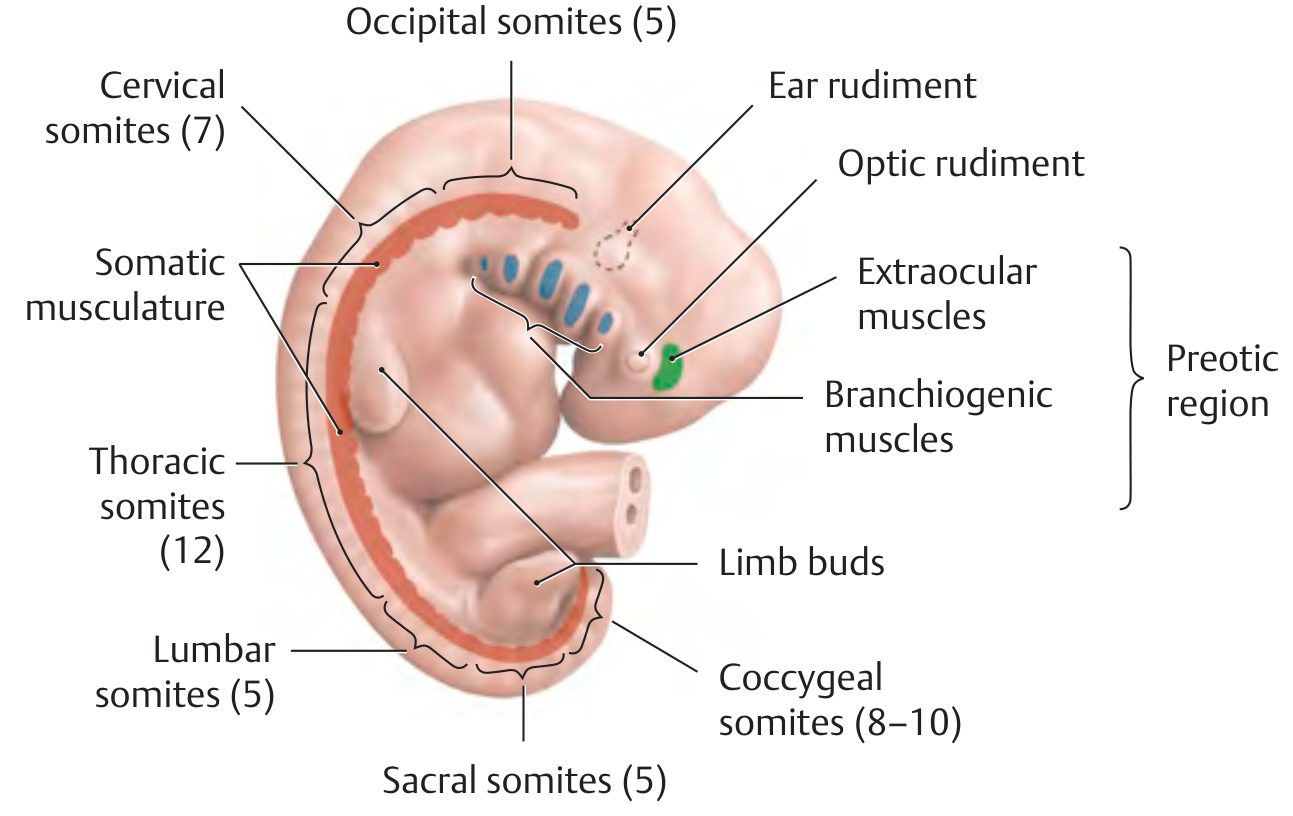
Before discrete somites form, the paraxial mesoderm in the head region first organizes into loosely arranged whorls called somitomeres (about 7 pairs in the cranial region). These somitomeres do not form discrete epithelial blocks and contribute to the branchiogenic pharyngeal arch muscles and extraocular muscles, innervated by cranial nerves.
From the occipital region caudally, somitomeres condense further into distinct paired blocks - the true somites.
3. Timeline and Numbers
Toward the end of the third week (day 20), the paraxial mesoderm begins segmenting into paired cuboidal blocks in a strict craniocaudal sequence:
| Period | Somite pairs |
|---|---|
| Day 20 | First pair appears (occipital region) |
| Days 26-32 | 38-39 pairs form (the "somite period") |
| End of week 5 | 42-44 pairs total |
Final distribution of somite pairs:
| Region | Pairs |
|---|---|
| Occipital | 4 (1st occipital disappears) |
| Cervical | 8 |
| Thoracic | 12 |
| Lumbar | 5 |
| Sacral | 5 |
| Coccygeal | 8-10 (most caudal 5-7 disappear) |
Because somites are so prominent and countable, somite number is used as a staging criterion for embryo age during weeks 4-5. - The Developing Human, Clinically Oriented Embryology
4. Molecular Control - The Segmentation Clock
Somite formation is not random; it is tightly regulated by a molecular oscillator (segmentation clock):
- WNT, FGF, and NOTCH signaling pathways interact in the presomitic mesoderm (PSM)
- Delta-Notch signaling controls the craniocaudal segmental pattern
- Tbx6 (T-box transcription factor) plays a key role in somitogenesis
- FoxC1 and FoxC2 (forkhead transcription factors) are expressed before somite formation begins
- HOX genes provide positional identity along the cranio-caudal axis
A traveling wave of gene expression sweeps caudally through the PSM; each cycle of the clock pinches off one new somite pair - approximately 3 pairs per day. - The Developing Human
5. Somite Structure
Each newly formed somite is initially a cuboidal epithelial block arranged around a small central cavity (the somitocoel). In transverse section, somites appear triangular and form visible ridges on the dorsal surface of the embryo.
6. Somite Differentiation - The Three Compartments
Once formed, each somite differentiates into two main regions, which give rise to three functional compartments:
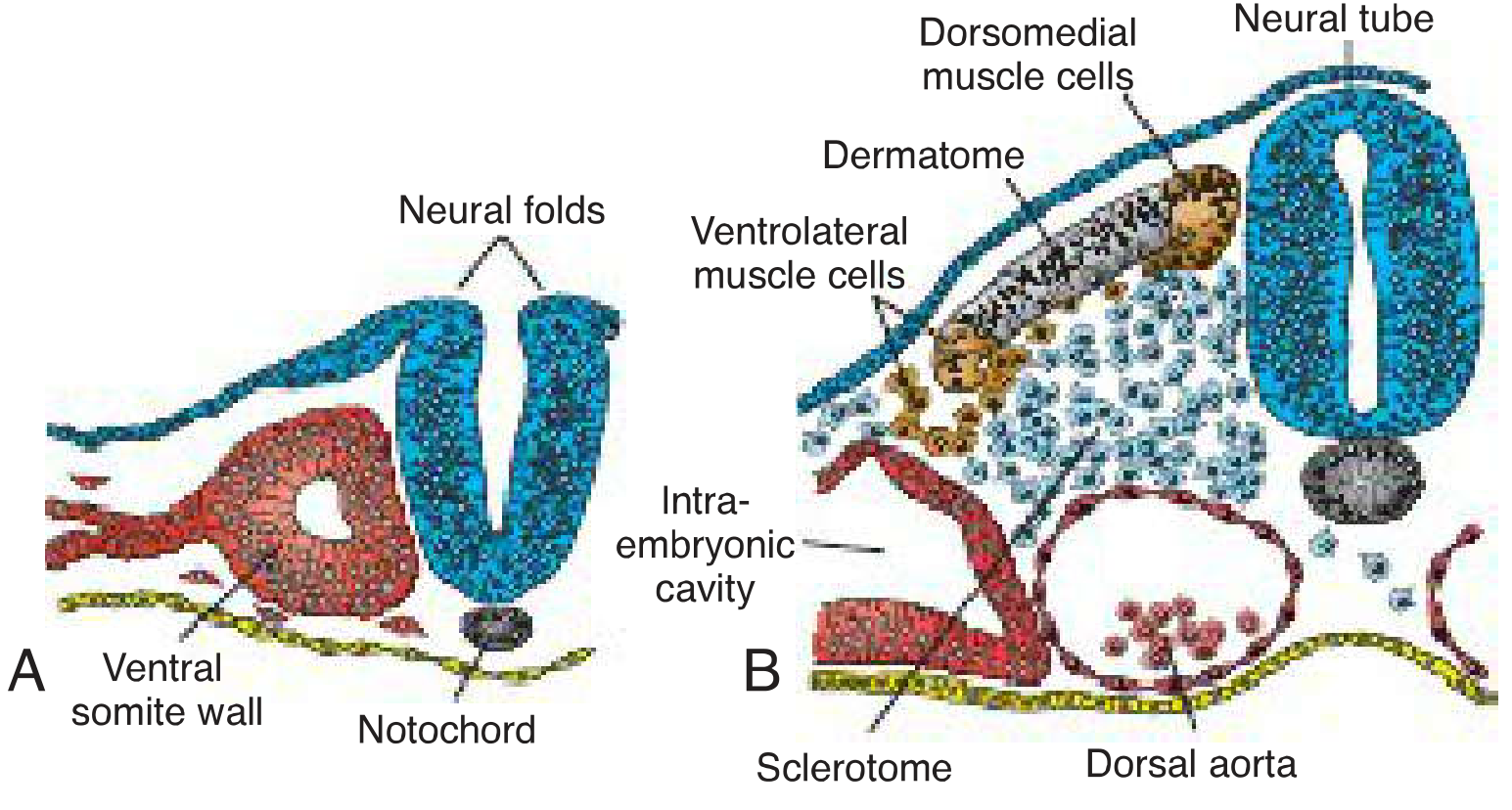
A. Sclerotome (ventromedial)
- Cells of the ventromedial wall lose their epithelial arrangement and become mesenchymal
- They migrate around the notochord and neural tube
- Differentiate into chondroblasts and osteoblasts forming the vertebrae, ribs, and intervertebral discs
- The notochord induces sclerotome formation via Sonic Hedgehog (SHH) signaling
B. Dermomyotome (dorsolateral)
The remaining dorsolateral epithelium = dermomyotome, which splits into:
Myotome (muscle-forming):
- Cells migrate beneath the dermatome to form two muscle masses:
- Epimere (epaxial myotome) - innervated by dorsal (posterior) rami of spinal nerves → intrinsic back muscles (e.g., erector spinae, rotatores, interspinales, intertransverse muscles)
- Hypomere (hypaxial myotome) - innervated by ventral (anterior) rami → anterolateral muscles of thorax/abdomen and limb musculature
- In limb bud regions, myoblasts migrate from the myotome into the limb bud and differentiate into dorsal (extensor) and ventral (flexor) muscle masses
Dermatome (dermis-forming):
- The remaining dorsal epithelial strip
- Cells migrate to the surface ectoderm of the corresponding segment and form the dermis of the skin (together with lateral plate mesoderm)
7. Signals Governing Differentiation
| Structure | Signal | Effect on somite |
|---|---|---|
| Notochord + floor plate | Sonic Hedgehog (SHH) | Induces sclerotome (ventral fate) |
| Dorsal neural tube | Wnt1/Wnt3a | Induces dermomyotome (dorsal fate) |
| Surface ectoderm | Wnt7a | Maintains/activates Myf5 in myotome |
| Notochord + floor plate (if transplanted dorsally) | SHH | Completely inhibits myotome; converts to cartilage instead |
8. Summary of Derivatives
| Somite Compartment | Derivatives |
|---|---|
| Sclerotome | Vertebrae, ribs, intervertebral discs |
| Myotome | All skeletal muscles of trunk and limbs |
| Dermatome | Dermis of the back and trunk |
The original segmental arrangement largely disappears with development - it persists only in the deep intrinsic back muscles (rotatores, interspinal, intertransverse) and the intercostal muscles. Superficial myotomes fuse into long, continuous muscles ("polymerization"), though the neurovascular supply still reflects the original segmental organization. - THIEME Atlas of Anatomy
Clinical note: Errors in somitogenesis (e.g., Notch signaling mutations) can lead to spondylocostal dysostosis (irregular vertebrae and rib fusions). Proper HOX gene expression is essential for correct vertebral identity - disruptions cause homeotic transformations (e.g., cervical ribs).
This is a shared conversation. Sign in to Orris to start your own chat.