Here are your complete, detailed study notes drawn directly from Marks' Basic Medical Biochemistry: A Clinical Approach, 6th Edition (Lieberman, Marks & Peet, 2022 - Wolters Kluwer), Chapter 39, pp. 665-671.
Synthesis & Degradation of Pyrimidine Nucleotides
Marks' Basic Medical Biochemistry, 6th Ed. — Chapter 39
Preliminary Concepts: Purines vs. Pyrimidines
Both purines and pyrimidines are required for synthesizing nucleotides and nucleic acids. They can be made de novo (from scratch) or salvaged from existing bases. Dietary intake of bases is low because intestinal epithelial cells metabolize most ingested nucleic acids before absorption.
Key difference in synthesis strategy:
- Purines are built on ribose 5-phosphate (ring assembled on the sugar first)
- Pyrimidines are synthesized as a free base first, then attached to ribose 5-phosphate
I. Overview: Origins of the Purine Ring (Context)
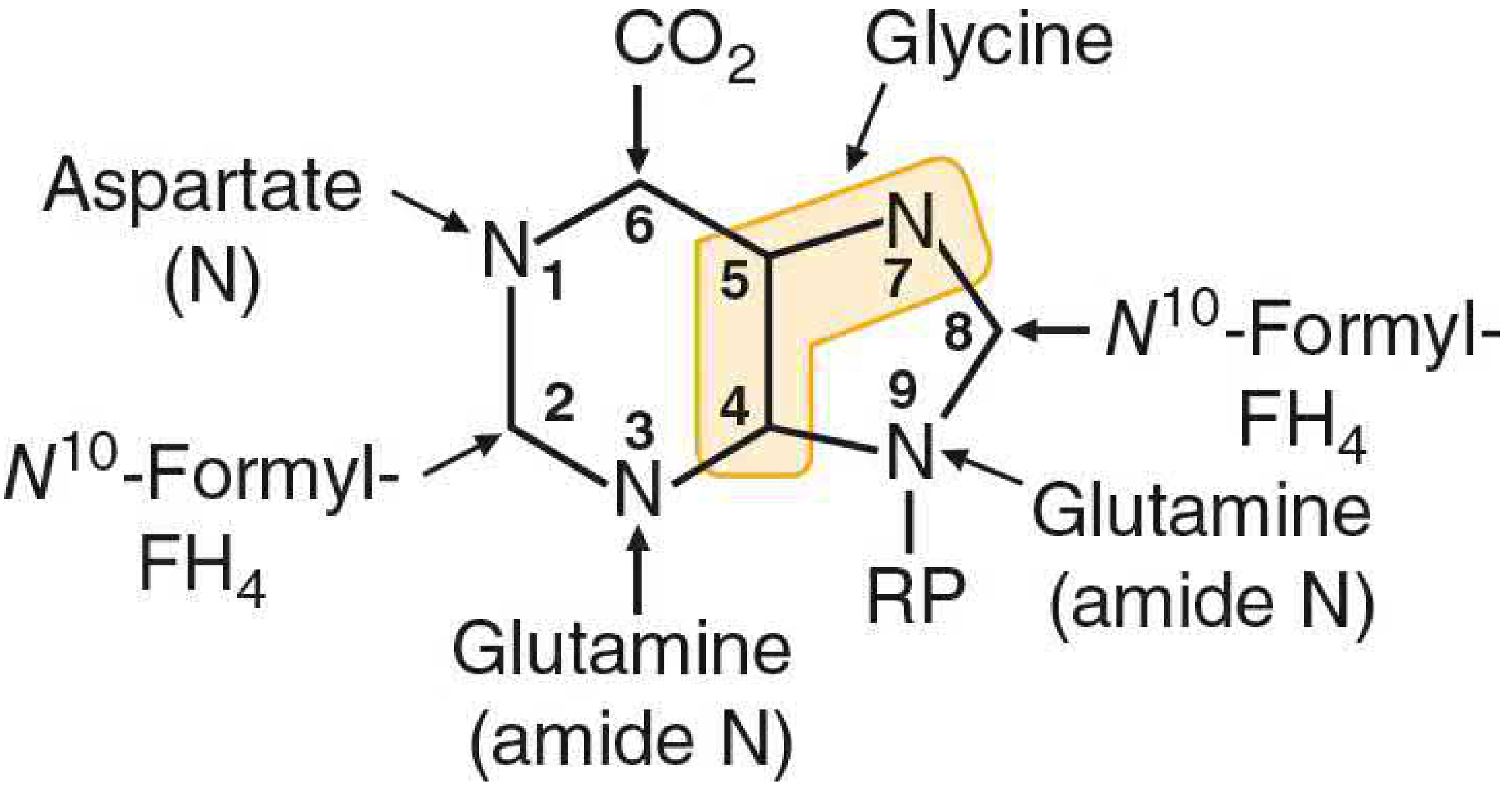
Figure 39.1 - Origin of Purine Ring Atoms:
The purine ring is assembled from: glycine (C4, C5, N7), CO2 (C6), aspartate nitrogen (N1), glutamine amide N (N3, N9), and two N10-formyl-FH4 units (C2, C8). The initial purine made is IMP, from which AMP and GMP are derived.
II. Synthesis of Pyrimidine Nucleotides
A. De Novo Pathway
The critical conceptual difference: In pyrimidine synthesis, the pyrimidine ring is synthesized as a free base first, then attached to ribose 5-phosphate. This is the opposite of purine synthesis.
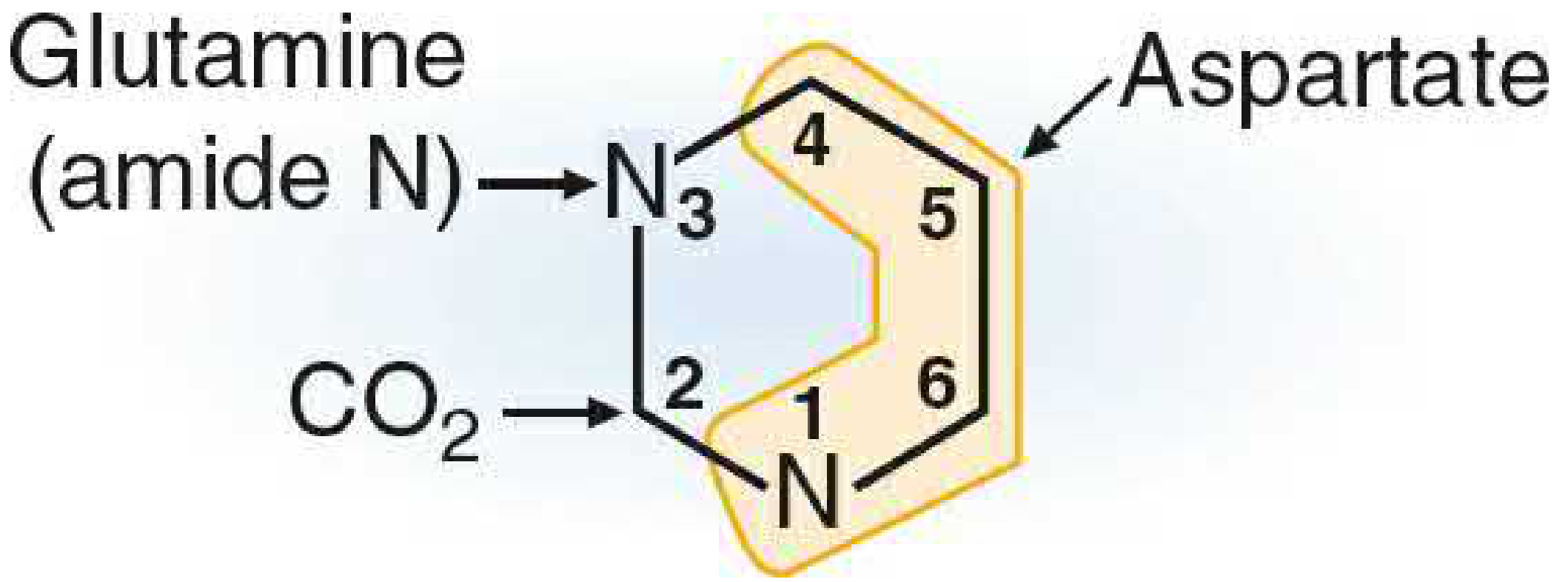
Origin of Pyrimidine Ring Atoms
Figure 39.14 - Origin of atoms in the pyrimidine ring:
| Ring Position | Contributed By |
|---|
| N-1 | Aspartate (nitrogen) |
| C-2 | CO2 |
| N-3 | Glutamine (amide nitrogen) |
| C-4, C-5, C-6 | Aspartate (carbon skeleton) |
In other words: Aspartate contributes the entire carbon backbone + N1; Glutamine amide N becomes N3; CO2 becomes C2.
Step-by-Step Pathway (6 reactions to UMP)
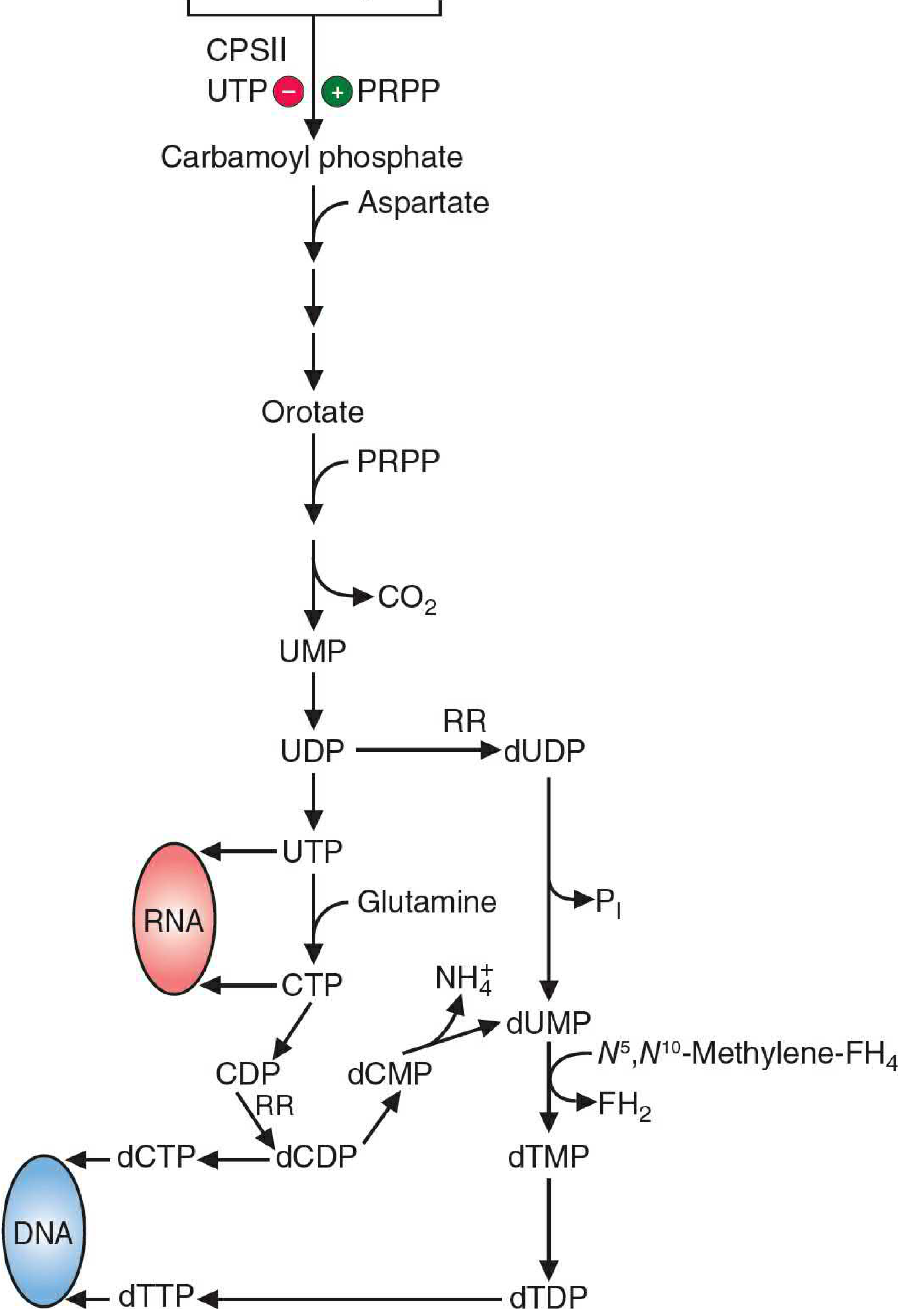
Figure 39.13 - Full pyrimidine synthesis pathway (overview):
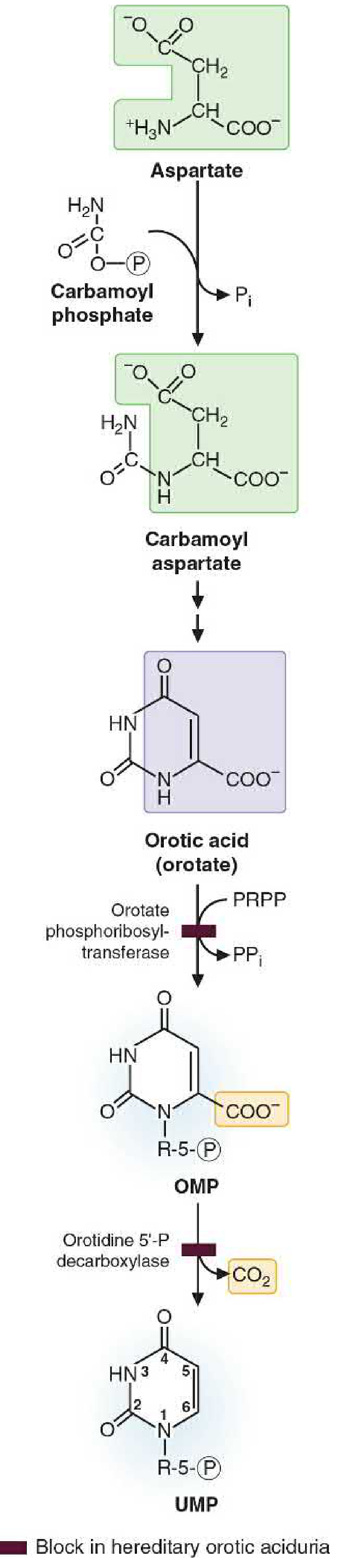
Figure 39.15 - Detailed conversion of carbamoyl phosphate + aspartate → UMP:
Step 1: Carbamoyl Phosphate Synthesis (cytosol)
Glutamine + CO2 + 2 ATP → Carbamoyl phosphate + Glutamate + 2 ADP + Pi
- Enzyme: Carbamoyl Phosphate Synthetase II (CPSII)
- Location: Cytosol (contrast with CPSI which is in mitochondria for the urea cycle)
- This is the regulated (committed) step of pyrimidine biosynthesis
- Nitrogen source is glutamine (not ammonia as in CPSI)
Critical Comparison — CPSI vs CPSII:
| Feature | CPSI (Urea Cycle) | CPSII (Pyrimidine Synthesis) |
|---|
| Pathway | Urea cycle | Pyrimidine biosynthesis |
| Nitrogen source | NH4+ (ammonia) | Glutamine (amide N) |
| Location | Mitochondria | Cytosol |
| Activator | N-acetylglutamate | PRPP |
| Inhibitor | None listed | UTP |
Step 2: Condensation with Aspartate
Carbamoyl phosphate + Aspartate → N-Carbamoyl aspartate + Pi
- Enzyme: Aspartate transcarbamoylase (ATCase)
- The entire aspartate molecule adds to carbamoyl phosphate
Step 3: Ring Closure
N-Carbamoyl aspartate → Dihydroorotate + H2O
- Enzyme: Dihydroorotase
- The linear molecule cyclizes to form the 6-membered pyrimidine ring (still in reduced form)
Step 4: Oxidation → Orotate
Dihydroorotate → Orotate (orotic acid)
- Enzyme: Dihydroorotate dehydrogenase
- Located on the outer surface of the inner mitochondrial membrane (the only mitochondrial step)
- Orotate = the first aromatic pyrimidine - it is the characteristic intermediate
Step 5: Attachment of Ribose 5-Phosphate
Orotate + PRPP → Orotidine 5'-monophosphate (OMP) + PPi
- Enzyme: Orotate phosphoribosyltransferase
- PRPP = 5'-phosphoribosyl-1-pyrophosphate (the activated form of ribose 5-phosphate)
- This is where the base gets linked to the sugar phosphate to become a nucleotide
Step 6: Decarboxylation → UMP
OMP → UMP + CO2
- Enzyme: Orotidine 5'-phosphate (OMP) decarboxylase (also called orotidylic acid decarboxylase)
- Product: Uridine Monophosphate (UMP) - the first pyrimidine nucleotide
Multifunctional Enzyme Complexes (Mammals)
A key feature of mammalian pyrimidine synthesis is that the enzymes are organized into bifunctional polypeptides:
- CAD protein (single polypeptide): contains CPSII + Aspartate transcarbamoylase + Dihydroorotase (steps 1-3)
- UMP Synthase (single polypeptide): contains Orotate phosphoribosyltransferase + OMP decarboxylase (steps 5-6)
Clinical relevance: In Hereditary Orotic Aciduria, the UMP synthase bifunctional enzyme is defective (see Clinical Correlation below).
From UMP to Other Pyrimidine Nucleotides
Following UMP formation:
- UMP → UDP → UTP (two sequential phosphorylation reactions by kinases)
- UTP → CTP: CTP synthetase transfers an amino group from glutamine amide N to carbon-4 of UTP
- This reaction requires ATP
- Cannot happen at the monophosphate level
- UTP and CTP are both precursors for RNA synthesis
- Deoxypyrimidines are produced via ribonucleotide reductase and thymidylate synthase (see Section IV)
III. Salvage of Pyrimidine Bases
Pyrimidines can be recycled from free bases produced during nucleotide degradation. The salvage pathway operates in two steps:
Step 1: Base → Nucleoside
Pyrimidine base + Ribose 1-phosphate → Nucleoside + Pi
- Enzyme: Pyrimidine nucleoside phosphorylase (relatively non-specific - works on multiple pyrimidines)
- Note: The preferred direction here is the reverse phosphorylase reaction (phosphate is released, not used as nucleophile). The base is added to the sugar.
- Substrates: uracil, cytosine, thymine
Step 2: Nucleoside → Nucleotide
Nucleoside + ATP → Nucleotide + ADP
- Enzymes: Nucleoside kinases (more substrate-specific than phosphorylase)
- Examples: Uridine kinase, Thymidine kinase (TK), Deoxycytidine kinase
Special mention - Thymidine Kinase (TK):
- Allosterically inhibited by dTTP (product feedback)
- Activity is tightly linked to the cell cycle: rises dramatically as cells enter S-phase
- Rapidly dividing cells (cancer, rapidly proliferating tissue) have high TK levels
- Radiolabeled thymidine is widely used for isotopic labeling of DNA and measuring intracellular DNA synthesis rates
Further phosphorylation: Additional kinases convert nucleoside monophosphates to diphosphates and triphosphates (NMP → NDP → NTP), the same pathway used for purines.
IV. Regulation of De Novo Pyrimidine Synthesis
The key regulated enzyme is CPSII (step 1 of the pathway).
Allosteric Regulation
| Effector | Effect on CPSII |
|---|
| UTP (end product) | Inhibits (feedback inhibition) |
| PRPP | Activates |
When pyrimidine levels drop (low UTP), CPSII is relieved of inhibition and synthesis accelerates. When UTP is abundant, CPSII is switched off.
Cell-Cycle Regulation (Phosphorylation State)
CPSII activity is also regulated by phosphorylation linked to the cell cycle:
-
Approaching S-phase:
- CPSII becomes more sensitive to PRPP activation
- Less sensitive to UTP inhibition
- This primes the cell for high nucleotide production needed for DNA replication
- Mechanism: phosphorylation by MAP kinase at a specific site → more easily activated enzyme
-
End of S-phase:
- UTP inhibition becomes more pronounced
- PRPP activation is reduced
- Mechanism: phosphorylation by cAMP-dependent protein kinase → more easily inhibited enzyme
This cell-cycle-linked regulation ensures nucleotide availability is precisely matched to DNA synthesis demand.
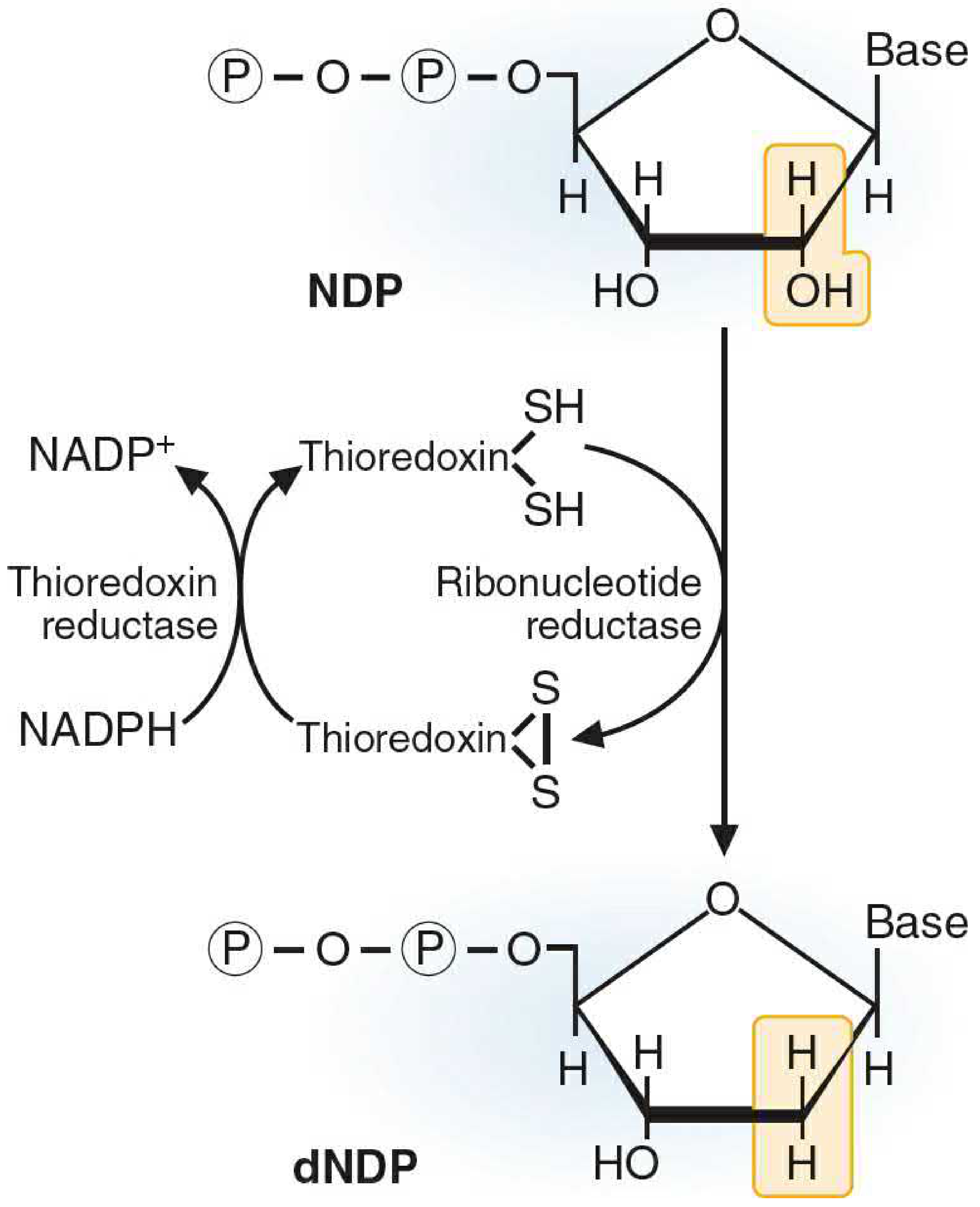
V. Production of Deoxyribonucleotides
For DNA synthesis, the ribose sugar must be reduced to 2'-deoxyribose. This cannot happen at the nucleoside level - it requires the diphosphate form.
Ribonucleotide Reductase
Figure 39.17 - Reduction of ribose to deoxyribose:
NDP → dNDP (reduction at the 2' position of ribose)
Substrates: ADP, GDP, CDP, UDP (all four nucleoside diphosphates)
Cofactor requirement: The reducing agent is the protein thioredoxin (which has two -SH groups). When thioredoxin donates its electrons to ribonucleotide reductase, it becomes oxidized to the disulfide form (-S-S-). Thioredoxin is re-reduced by thioredoxin reductase using NADPH.
After reduction: The dNDPs are phosphorylated to dNTPs (deoxyribonucleoside triphosphates) and used as precursors for DNA synthesis.
Allosteric Regulation of Ribonucleotide Reductase
The regulation is extremely complex. The enzyme has two allosteric sites:
- Overall Activity Site - controls whether the enzyme is ON or OFF
- Substrate Specificity Site - determines which NDP is preferentially reduced
Overall Activity Site:
- ATP bound → enzyme active
- dATP bound → enzyme inhibited (overall activity shut off)
Substrate Specificity Site (sequential control):
| Nucleotide at Specificity Site | Preferred Substrate Reduced |
|---|
| ATP or dATP | CDP and UDP (pyrimidines) |
| dTTP | GDP |
| dGTP | ADP |
| dATP (at activity site) | Enzyme OFF (all substrates) |
This creates an elegant sequential self-regulating system:
- ATP activates reduction of CDP and UDP → produces dCDP and dUDP
- dUDP → dUMP → dTMP → dTTP accumulates
- dTTP at specificity site → switches to reducing GDP → produces dGDP → dGTP
- dGTP at specificity site → switches to reducing ADP → produces dADP → dATP
- dATP at activity site → shuts down entire enzyme
This ensures balanced dNTP pools for DNA replication.
Table 39.3 - Summary of Ribonucleotide Reductase Effectors:
| Preferred Substrate | Activity Site Effector | Specificity Site Effector |
|---|
| None (OFF) | dATP | Any nucleotide |
| CDP | ATP | ATP or dATP |
| UDP | ATP | ATP or dATP |
| GDP | ATP | dTTP |
| ADP | ATP | dGTP |
dTMP Synthesis (Thymidylate Synthase)
dUMP (from dUDP dephosphorylation OR from dCMP deamination) is converted to dTMP by thymidylate synthase:
dUMP + N5,N10-Methylene-FH4 → dTMP + FH2 (dihydrofolate)
- The methyl group donor is N5,N10-methylene-tetrahydrofolate
- FH4 is oxidized to FH2 (dihydrofolate) in this reaction
- FH2 is regenerated to FH4 by dihydrofolate reductase (DHFR)
- Methotrexate (anticancer drug) and trimethoprim (antibiotic) inhibit DHFR, blocking this regeneration and halting DNA synthesis in rapidly dividing cells
VI. Degradation of Purine and Pyrimidine Bases
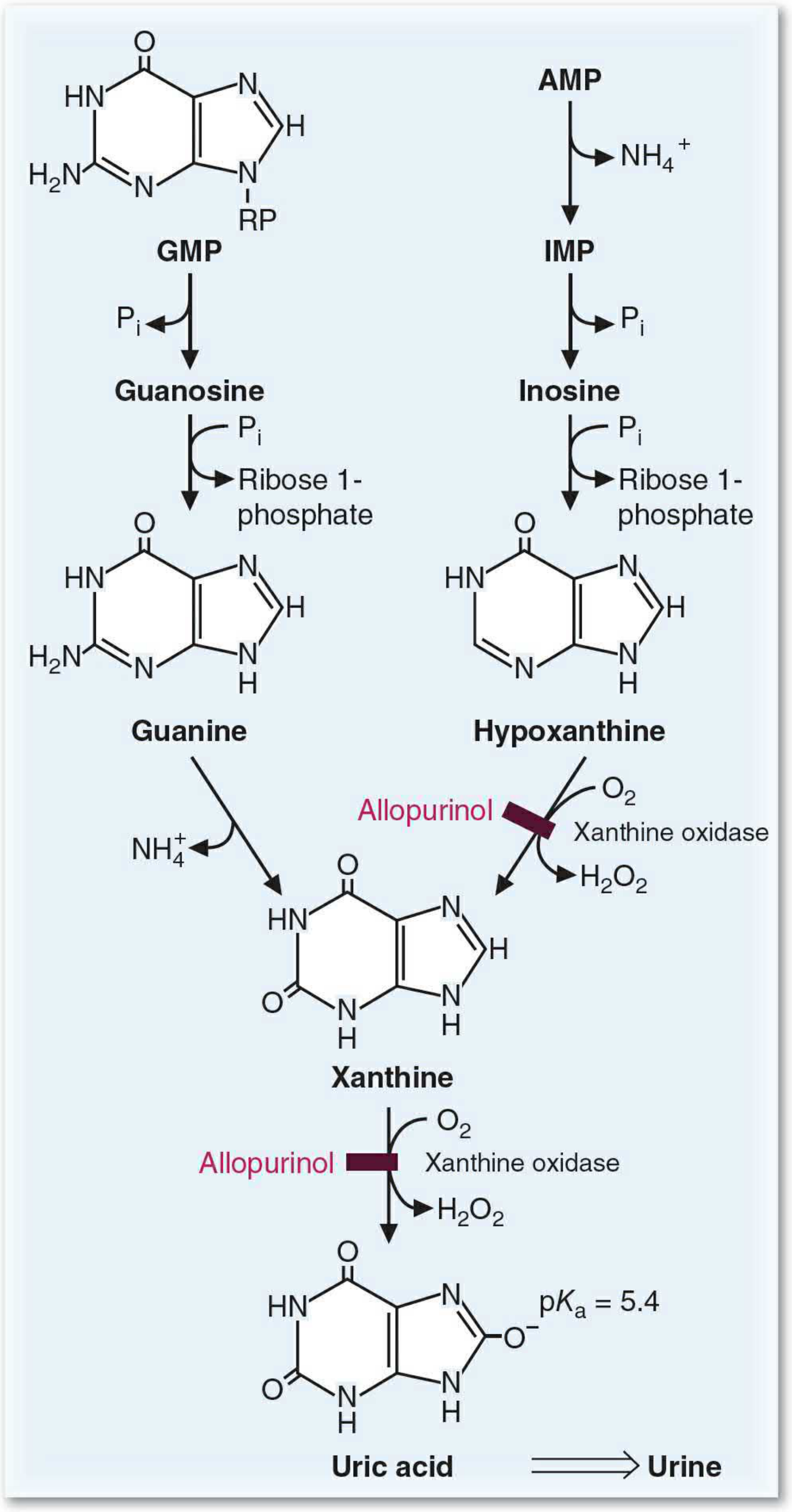
A. Degradation of Purine Bases
Figure 39.18 - Degradation of purine bases to uric acid:
Degradation of purine nucleotides occurs mainly in the liver. The salvage enzymes handle most reactions:
Pathway:
- AMP → deaminated (AMP deaminase) → IMP + NH4+
- IMP and GMP → dephosphorylated (5'-nucleotidase) → inosine and guanosine
- Inosine and guanosine → cleaved by purine nucleoside phosphorylase → free bases + ribose 1-phosphate
- Inosine → hypoxanthine
- Guanosine → guanosine → deaminated by guanosine → guanine → deamination → xanthine
- Hypoxanthine → oxidized by xanthine oxidase → xanthine
- Xanthine → oxidized by xanthine oxidase → Uric acid
Pathways for adenine and guanine converge at xanthine and end at uric acid.
Xanthine Oxidase:
- Molybdenum-requiring enzyme
- Uses molecular oxygen (O2) as electron acceptor
- Produces H2O2 as a byproduct
- A second form uses NAD+ as the electron acceptor instead
- Allopurinol (used to treat gout) is a structural analog of hypoxanthine that inhibits xanthine oxidase
Uric Acid:
- pKa = 5.4, so it exists as urate (ionized) in the body
- Urate is poorly soluble in aqueous environments
- Normal blood urate concentration is close to the solubility limit
- Gout = accumulation of urate crystals in joints (especially the big toe) due to hyperuricemia
- Very little energy is recovered from purine ring degradation - this is why salvage is metabolically preferred
Clinical Note: To determine whether a gout patient overproduces purines (vs. underexcretes), an oral dose of 15N-labeled glycine can be given - glycine's entire nitrogen appears in the purine ring and subsequently in uric acid, allowing measurement of new purine synthesis.
B. Degradation of Pyrimidine Bases
Pyrimidine degradation is simpler and produces water-soluble, non-toxic end-products (unlike purines which produce problematic urate):
Pathway:
- Pyrimidine nucleotides → dephosphorylated → nucleosides
- Nucleosides → cleaved → ribose 1-phosphate + free pyrimidine bases (cytosine, uracil, thymine)
- Cytosine → deaminated → Uracil
- Uracil → degraded → CO2 + NH4+ + β-alanine
- Thymine → degraded → CO2 + NH4+ + β-aminoisobutyrate
End-products: β-alanine, β-aminoisobutyrate, CO2, NH4+
- These are either excreted in urine or converted to CO2, H2O, and NH4+ (which enters urea synthesis)
- No problematic precipitating product - this is why pyrimidine disorders do not cause gout-like symptoms
- Like purines, little energy is recovered from pyrimidine ring degradation
VII. Clinical Correlation: Hereditary Orotic Aciduria
Molecular Basis
Hereditary orotic aciduria is caused by a genetic defect in UMP synthase, the bifunctional enzyme that carries out the last two steps of de novo pyrimidine synthesis:
- Orotate phosphoribosyltransferase (step 5: orotate + PRPP → OMP)
- OMP decarboxylase (step 6: OMP → UMP + CO2)
Both activities reside on the same polypeptide (UMP synthase). A loss-of-function mutation in either activity blocks the pathway at orotate, causing orotate to accumulate.
The block is shown in Figure 39.15 by the dark bar - indicating where the pathway is blocked in this condition.
Consequences
- Orotate (orotic acid) cannot be converted to UMP → it accumulates and is excreted in urine (orotic aciduria)
- UMP cannot be synthesized de novo
- Without UMP, cells cannot make UTP, CTP, dCTP, or dTMP
- Normal growth does not occur - children present with growth retardation/failure to thrive and megaloblastic anemia (cells cannot divide because they lack nucleotides for DNA synthesis)
Treatment
Oral administration of uridine (a nucleoside, not the base)
Uridine enters cells and is converted directly to UMP by uridine kinase, bypassing the metabolic block entirely. From UMP, the body can then synthesize:
- UTP (via phosphorylation)
- CTP (via CTP synthetase)
- dTMP (via ribonucleotide reductase and thymidylate synthase)
This treatment fully corrects the pyrimidine deficiency.
Important Differential: OTC Deficiency vs. Hereditary Orotic Aciduria
Both conditions cause orotic aciduria but by entirely different mechanisms:
| Feature | Hereditary Orotic Aciduria | OTC Deficiency (Ornithine Transcarbamoylase) |
|---|
| Cause | Defect in UMP synthase | Defect in urea cycle enzyme (OTC) |
| Mechanism | Cannot convert orotate → UMP | Excess carbamoyl phosphate leaks from mitochondria into cytosol, bypasses CPSII regulation, floods pyrimidine pathway |
| Ammonia levels | Normal | Elevated (hyperammonemia) |
| Growth | Retarded | Hyperammonemia symptoms dominate |
| Treatment | Uridine supplementation | Protein restriction + nitrogen scavenging agents |
In OTC deficiency, mitochondrial carbamoyl phosphate accumulates (because it cannot enter the urea cycle), leaks into the cytoplasm, and bypasses the regulated CPSII step - flooding the pyrimidine synthesis pathway and producing excess orotate. The result is orotic aciduria without pyrimidine deficiency.
VIII. Summary Table: Key Diseases in Nucleotide Metabolism (Table 39.4)
| Disease | Type | Key Mechanism |
|---|
| Gout | Both (genetic + environmental) | Elevated urate → crystal deposition in joints; painful arthritis |
| Hereditary Orotic Aciduria | Genetic | Defect in UMP synthase → orotic acid accumulation + growth failure |
| Lesch-Nyhan Syndrome | Genetic | Loss of HGPRT → purines not salvaged → uric acid overproduction + intellectual disability + self-injury |
| PNP Deficiency | Genetic | Defect in purine nucleoside phosphorylase → T-cell dysfunction → partial immunodeficiency |
| ADA Deficiency | Genetic | Loss of adenosine deaminase → deoxyadenosine/dATP accumulation → SCID (T + B cell loss) |
IX. Key Concepts Summary (for Exam)
-
Pyrimidine ring is made from aspartate + carbamoyl phosphate (derived from CO2 + glutamine). No formyl-FH4 needed (unlike purines).
-
Order of pyrimidine synthesis: Base is made FIRST → then attached to PRPP. Sequence: Carbamoyl-P → Carbamoyl aspartate → Dihydroorotate → Orotate → OMP → UMP → UDP → UTP → CTP.
-
Regulated step = CPSII: Inhibited by UTP (end product feedback), activated by PRPP.
-
CPSII vs CPSI: Same carbon source (CO2), different nitrogen source (glutamine vs. NH4+), different location (cytosol vs. mitochondria), different allosteric regulators.
-
Salvage of pyrimidines: Base → nucleoside (pyrimidine nucleoside phosphorylase) → nucleotide (nucleoside kinase).
-
Thymidine kinase: Rises in S-phase; used clinically to assess cell proliferation; inhibited by dTTP.
-
Ribonucleotide reductase makes all four deoxyribonucleotides; regulated by two allosteric sites (activity + specificity); requires thioredoxin + NADPH.
-
dTMP synthesis: dUMP + N5,N10-methylene-FH4 → dTMP + FH2. Target of methotrexate (inhibits DHFR).
-
Purine degradation endpoint = uric acid; insoluble; excreted in urine. Gout from hyperuricemia. Allopurinol inhibits xanthine oxidase.
-
Pyrimidine degradation endpoint = β-alanine + β-aminoisobutyrate + CO2 + NH4+; water-soluble, non-toxic, excreted easily.
-
Hereditary orotic aciduria: UMP synthase defect → cannot convert orotate → UMP. Treat with uridine orally. Normal ammonia levels (not hyperammonemia).
All content sourced directly from: Marks' Basic Medical Biochemistry: A Clinical Approach, 6th Ed. (Lieberman, Marks & Peet, 2022, Wolters Kluwer), Chapter 39, pp. 665-671 (book pp. 1406-1436).