fully detailed and structured notes on muslce spindle and golgi tendon apparatus ( write about the structre,receptor function of muscle spindle, muscle spindle reflex, role of the muscle spindle in voluntary motor activity, golgi tendon reflex, etc using Guyton and hall and ganong etc books as your source)
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Reading File
Loading Image
Loading Image
I now have comprehensive, authoritative content from Guyton & Hall, Kandel's Principles of Neural Science, and Costanzo Physiology. Let me now compose the full structured notes:
Muscle Spindle and Golgi Tendon Apparatus - Detailed Structured Notes
Primary Sources: Guyton & Hall Textbook of Medical Physiology (14th ed.); Kandel: Principles of Neural Science (6th ed.); Costanzo Physiology (7th ed.)
PART I: THE MUSCLE SPINDLE
1. Overview and Location
The muscle spindle is the principal sensory receptor for detecting changes in muscle length and the rate of change of that length. It is a small encapsulated structure embedded within skeletal muscle, oriented parallel to the surrounding large (extrafusal) muscle fibers. Each spindle is 3-10 mm long and is built around 3-12 tiny intrafusal muscle fibers whose ends attach to the glycocalyx of the surrounding extrafusal fibers.
(Guyton & Hall, p. 677)
2. Structure of the Muscle Spindle

Fig. 55.3 - Guyton & Hall: Muscle spindle showing its relationship to large extrafusal skeletal muscle fibers, with both motor and sensory innervation labeled.
A. Intrafusal Muscle Fibers (two types)
There are two distinct populations of intrafusal fibers within each spindle:
| Feature | Nuclear Bag Fibers | Nuclear Chain Fibers |
|---|---|---|
| Number per spindle | 1-3 | 3-9 |
| Diameter/length | Larger, longer | ~half the diameter and length |
| Nuclear arrangement | Nuclei clustered in a swollen "bag" in the central region | Nuclei aligned in a single chain throughout the receptor area |
| Primary function | Detects dynamic changes (velocity of stretch) | Detects static changes (sustained length) |
| Motor innervation | Dynamic gamma (gamma-d) fibers - via "plate endings" | Static gamma (gamma-s) fibers - via "trail endings" |
The central (equatorial) region of each intrafusal fiber contains few or no actin/myosin filaments and therefore cannot contract - it serves purely as the sensory receptor zone. The polar (end) regions contain contractile filaments and are activated by gamma motor neurons.
(Guyton & Hall, p. 677)
B. Sensory (Afferent) Innervation
Two types of sensory endings are located in the central receptor area:
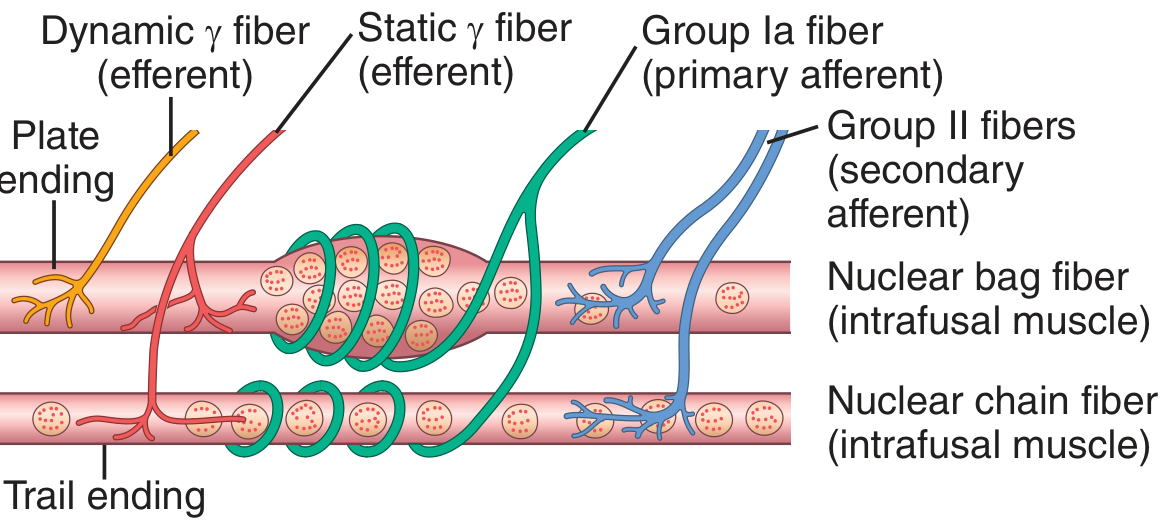
Fig. 55.4 - Guyton & Hall: Details of nerve connections from nuclear bag and nuclear chain muscle spindle fibers.
1. Primary Afferent Ending (Annulospiral Ending)
- Large Type Ia fiber (average diameter: 17 micrometers)
- Wraps spirally around the central portion of both nuclear bag and nuclear chain fibers
- Conduction velocity: 70-120 m/sec - among the fastest in the entire body
- Detects both the dynamic (velocity-sensitive) response and the static (length) response
- One primary ending per spindle
2. Secondary Afferent Ending (Flower Spray Ending)
- Smaller Type II fiber (average diameter: 8 micrometers)
- Located on one or both sides of the primary ending
- Innervates primarily the nuclear chain fibers
- Detects mainly the static (length) response - signals sustained stretch
- Usually 1, sometimes 2 secondary endings per spindle
(Guyton & Hall, p. 677)
C. Motor (Efferent) Innervation
The spindle receives motor innervation from gamma motor neurons in the anterior horn of the spinal cord, which are separate and distinct from the alpha motor neurons that drive extrafusal fibers.
- Gamma motor neurons are about half as numerous as alpha motor neurons
- Their axons (type A-gamma fibers) average 5 micrometers in diameter
- Two subtypes:
- Gamma-dynamic (gamma-d) neurons - innervate nuclear bag fibers - dramatically enhance the dynamic response of the spindle
- Gamma-static (gamma-s) neurons - innervate nuclear chain fibers - enhance the static response of the spindle
- Alpha motor neurons (type Aa fibers) average 14 micrometers and innervate extrafusal muscle
(Guyton & Hall, p. 676)
3. Receptor Function of the Muscle Spindle
The spindle receptor can be excited in two distinct ways:
- Passive stretch of the whole muscle - stretches the mid-portion of the spindle and excites the receptor
- Active contraction of the intrafusal fiber ends - even without change in whole muscle length, contraction of the polar regions stretches the non-contractile central zone, exciting the receptor
A. Dynamic Response
- Generated mainly by the primary (Ia) endings on nuclear bag fibers
- Sensitive to the rate of change of muscle length (velocity)
- When a muscle is suddenly stretched, the primary ending fires at a very high rate proportional to the speed of stretch
- When stretch stops, firing rate rapidly drops back to the static level
- When the muscle shortens, firing rate decreases below baseline (negative signal)
- The gamma-d fiber system controls the gain (intensity) of this dynamic response
B. Static Response
- Generated by both primary (Ia) and secondary (II) endings on nuclear chain fibers
- Sensitive to the maintained length of the muscle
- Firing rate remains elevated for the entire duration of stretch, proportional to the degree of stretch
- Persists as long as the muscle stays lengthened
- The gamma-s fiber system controls the gain of the static response
C. Continuous Baseline Discharge
Under normal conditions, with some tonic level of gamma neuron activity, muscle spindles emit sensory impulses continuously at a low, resting rate. Stretching the spindle increases firing; shortening decreases firing. This bidirectional signaling allows the spindle to transmit both positive (increased stretch) and negative (decreased stretch) signals to the spinal cord at all times.
(Guyton & Hall, p. 678)
4. The Muscle Spindle Reflex (Stretch Reflex / Myotatic Reflex)
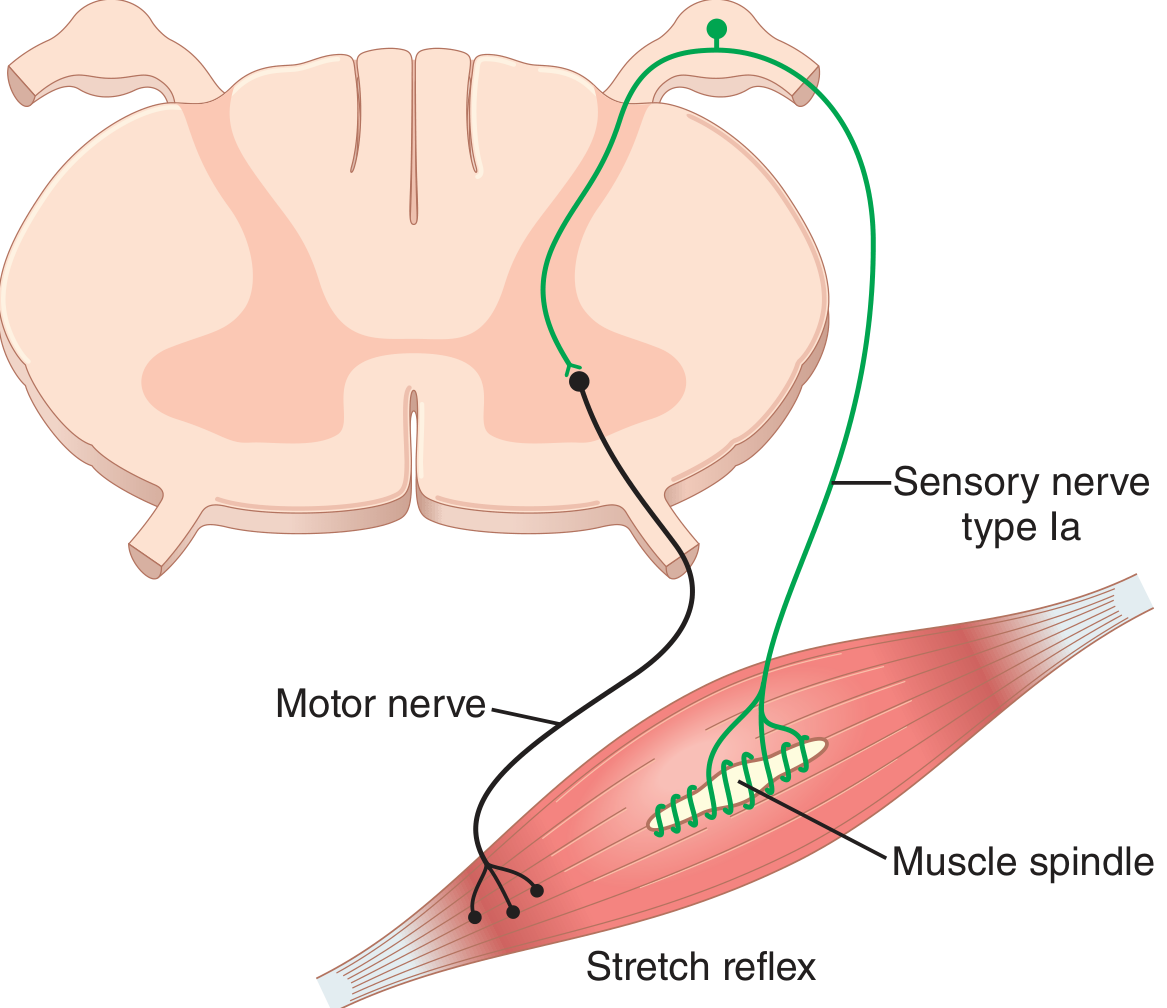
Fig. 55.5 - Guyton & Hall: Neuronal circuit of the stretch reflex.
A. Definition and Trigger
The myotatic reflex is the simplest and most well-characterized spinal reflex. Whenever a muscle is suddenly stretched, excitation of the spindles causes reflex contraction of the stretched muscle and its synergists.
Classic clinical test: the knee jerk (patellar reflex) - tapping the patellar tendon stretches the quadriceps, triggering its spindles, causing reflex quadriceps contraction.
B. Neural Circuit
The stretch reflex is primarily monosynaptic:
- Muscle is stretched
- Spindle intrafusal fiber midportion is deformed
- Ia (and II) afferent fibers fire
- Ia fiber enters the spinal cord via the dorsal root
- Ia fiber synapses directly on the alpha motor neuron in the anterior horn (monosynaptic connection - no interneuron for the Ia-alpha pathway)
- Alpha motor neuron fires, sending impulse back to the same muscle (homonymous muscle) via the ventral root
- Extrafusal fibers contract, opposing the stretch
- Type II fibers synapse via interneurons (polysynaptic), producing additional, delayed effects
The monosynaptic design minimizes the reflex time, making the response nearly instantaneous.
C. Reciprocal Inhibition (Reciprocal Innervation)
Simultaneously with excitation of the stretched muscle, Ia afferents activate Ia inhibitory interneurons that inhibit the antagonist muscle's alpha motor neurons. This reciprocal inhibition:
- Allows the agonist to contract without working against a tense antagonist
- Is a fundamental principle of spinal motor organization
D. Dynamic Stretch Reflex vs. Static Stretch Reflex
| Feature | Dynamic Stretch Reflex | Static Stretch Reflex |
|---|---|---|
| Signal source | Primary (Ia) endings on nuclear bag fibers | Primary (Ia) + Secondary (II) endings on nuclear chain fibers |
| Stimulus | Rapid change in muscle length | Sustained change in muscle length |
| Duration | Brief - over within fraction of a second | Prolonged - persists as long as stretch maintained |
| Magnitude | Strong, instantaneous | Weaker, sustained |
| Function | Opposes sudden changes in length | Maintains constant muscle length |
(Guyton & Hall, p. 679)
E. Damping/"Signal Averaging" Function
A crucial role of the stretch reflex is to smooth out jerky motor commands from higher brain centers. When the muscle spindle reflex is intact, even choppy supraspinal signals at 8 impulses/second produce smooth muscle contractions. After spindle sensory denervation, the same commands produce jerky, uncoordinated contractions (demonstrated in Guyton Fig. 55.6). This "signal averaging" or damping function smooths the translation of descending commands into graded, precise movements.
(Guyton & Hall, p. 679)
F. Clonus
When the stretch reflex is highly sensitized (as in spasticity or upper motor neuron lesions), a rhythmic oscillating contraction called clonus can result. It occurs when a sudden, sustained stretch sets up an oscillating cycle where the reflex contracts the muscle, which then releases the stretch, which then reduces the reflex, and the muscle stretches again. Testing for clonus is a neurological tool to assess the degree of supraspinal facilitatory drive on spinal circuits.
(Guyton & Hall, p. 681)
5. Role of the Muscle Spindle in Voluntary Motor Activity
A. Alpha-Gamma Coactivation
Approximately 31% of all motor fibers to muscle are small type A-gamma efferents. When the motor cortex or any brain area activates alpha motor neurons to produce voluntary movement, it simultaneously activates gamma motor neurons - a process called alpha-gamma coactivation.
Why coactivation is necessary:
-
Prevents spindle unloading: When extrafusal muscle contracts and shortens, the spindle (which lies in parallel) would passively shorten and go silent, removing all spindle-based reflex stabilization. Coactivation causes the intrafusal fiber ends to contract simultaneously, maintaining tension in the spindle's central region throughout the movement.
-
Maintains damping function: Continuous gamma drive keeps the spindle in its optimal operating range (neither too slack nor overstretched), ensuring it can continuously perform its damping and length-stabilizing roles regardless of the current muscle length.
(Guyton & Hall, p. 680)
B. Brain Centers Controlling the Gamma System
The gamma efferent system is controlled by:
- Bulboreticular facilitatory region (primary controller)
- Cerebellum (provides input to bulboreticular area)
- Basal ganglia (input to bulboreticular area)
- Cerebral cortex (input to bulboreticular area)
The bulboreticular facilitatory area is especially important for antigravity contractions, and antigravity muscles (e.g., extensors) have an exceptionally high density of muscle spindles. This suggests the gamma-spindle mechanism is particularly important for stabilizing posture and damping movements during locomotion (walking, running).
(Guyton & Hall, p. 680)
C. Stabilization of Body Position
During tense motor actions, the bulboreticular facilitatory region and associated brain areas greatly increase gamma motor neuron firing in all muscles simultaneously. This stiffens the spindle feedback loops throughout the body, increasing resistance of individual muscle groups to unexpected perturbations. The result is better postural stability during voluntary activity - the body becomes more resistant to being thrown off-balance by unexpected loads.
(Guyton & Hall, p. 680)
D. The Gamma Motor Loop ("Servo-Assist" Mechanism)
Some voluntary movements may be initiated or assisted partly through gamma activation first:
- Brain activates gamma neurons to contract intrafusal fiber ends
- This stretches the spindle's central region
- Ia signals fire, reflexively activating alpha motor neurons
- Extrafusal muscle contracts
This "servo-assist" or follow-up servo mechanism means the spindle system can help execute motor commands smoothly even when the alpha motor neuron drive is insufficient on its own.
6. Muscle Spindle and Fusimotor Control (from Kandel)
The fusimotor system's activity varies significantly with behavior type:
- Static gamma neurons are active during slow, predictable movements (where muscle length changes gradually)
- Dynamic gamma neurons are activated during fast, unpredictable movements (where length can change rapidly)
This differential activation allows the CNS to tune spindle sensitivity based on the type of movement expected - increasing dynamic sensitivity during rapid tasks to maintain accurate velocity feedback.
(Kandel, Principles of Neural Science, 6th ed., p. 817)
PART II: THE GOLGI TENDON APPARATUS
7. Structure of the Golgi Tendon Organ (GTO)

Fig. 55.8 - Guyton & Hall: Golgi tendon reflex. Excessive tension stimulates GTO receptors, signals travel via afferent fiber to inhibitory interneuron, inhibiting alpha motor neuron and causing muscle relaxation.
- Located at the musculotendinous junction (junction of muscle belly and tendon)
- Each organ is approximately 1 mm long and 0.1 mm in diameter
- Slender, encapsulated structure through which 10-15 tendon collagen fiber bundles pass
- Each collagen bundle is connected in series with a small group of extrafusal muscle fibers
- The capsule encloses braided collagen fascicles intertwined with the terminals of the sensory nerve
Note on arrangement: The GTO is arranged in series with muscle fibers (contrast with muscle spindles, which are in parallel). This series arrangement makes the GTO exquisitely sensitive to muscle tension (force), particularly that generated by active contraction.
(Kandel, Principles of Neural Science, 6th ed., p. 818)
Mechanism of GTO Activation
- When tension develops in the muscle (via contraction or passive stretch), the tendon fibers are stretched
- This straightens/tightens the braided collagen fibers within the capsule
- The collagen fibers compress the Ib nerve endings intertwined among them
- This mechanical compression is transduced into action potentials in the Ib afferent fiber
- Even very small stretches can compress the nerve endings because of the close anatomical association of nerve and collagen
- The GTO is most potently activated by active muscle contraction (which both shortens muscle and tenses the tendon)
(Kandel, Principles of Neural Science, 6th ed., p. 818)
8. Receptor Function of the Golgi Tendon Organ
A. Sensory Afferent Fiber (Group Ib)
- A single Ib afferent axon innervates each GTO
- Average diameter: 16 micrometers (only slightly smaller than Ia from muscle spindles)
- Conduction velocity: fast, large myelinated fiber
- The Ib axon branches extensively within the capsule
B. What the GTO Detects
The primary stimulus for GTO activation is muscle tension (force) - specifically, the tension transmitted through the small bundle of muscle fibers connected to that organ. This is the fundamental difference from the muscle spindle:
| Property | Muscle Spindle | Golgi Tendon Organ |
|---|---|---|
| Detects | Muscle length and rate of length change | Muscle tension (force) |
| Arrangement | Parallel to extrafusal fibers | Series with extrafusal fibers |
| Best stimulus | Passive stretch | Active muscle contraction |
| Afferent fiber | Ia (17 µm) and II (8 µm) | Ib (16 µm) |
| Efferent (motor) innervation | Gamma neurons (intrafusal) | None |
(Guyton & Hall, p. 681)
C. Dynamic and Static Responses
Like the muscle spindle's primary receptor, the GTO displays:
- Dynamic response: intense firing when muscle tension suddenly increases
- Static response: steady-state firing rate that is nearly directly proportional to muscle tension - settles within a fraction of a second
This means the GTO provides the nervous system with both instantaneous (dynamic) and continuous (static) information about the precise degree of tension in each small segment of each muscle.
D. Sensitivity
Contrary to early beliefs that GTOs were only "overload sensors," we now know they signal minute changes in muscle tension during normal, low-intensity contractions and movements. Single Ib axon recordings in humans during voluntary finger movements and in cats during normal walking confirm that GTOs are active during everyday movements, not just extreme loading situations.
(Kandel, Principles of Neural Science, 6th ed., pp. 817-818)
9. The Golgi Tendon Reflex (Inverse Myotatic Reflex / Autogenic Inhibition)
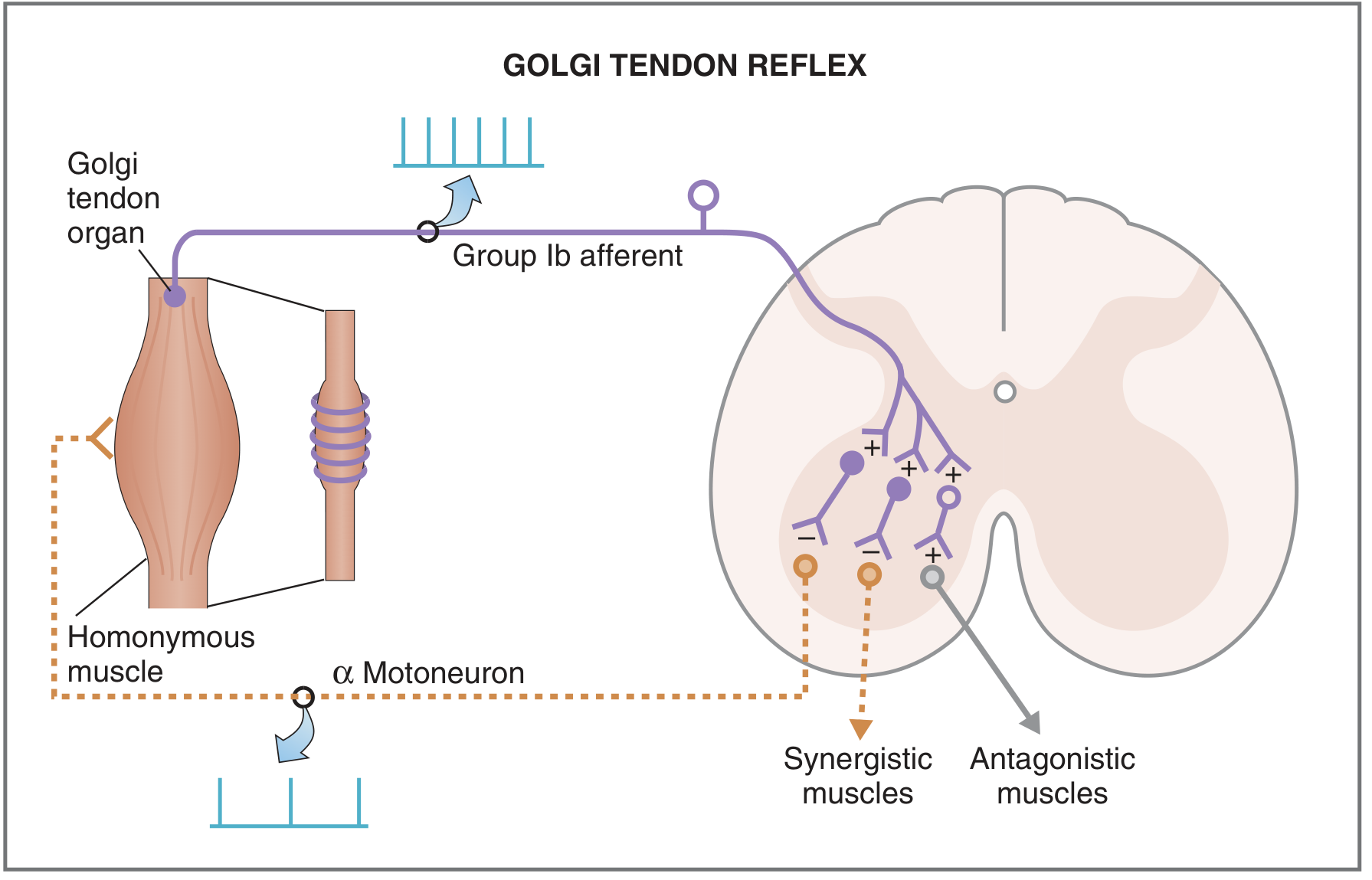
Fig. 3.34 - Costanzo: Golgi tendon reflex pathway. Solid lines = excitatory pathways; dashed lines = inhibitory steps.
A. Definition
The Golgi tendon reflex is a disynaptic (two-synapse) spinal cord reflex also called:
- The inverse myotatic reflex (because it produces effects opposite to the stretch reflex)
- Autogenic inhibition (because stimulation of a muscle's own tension receptors inhibits that same muscle)
(Costanzo Physiology, 7th ed., p. 111)
B. Reflex Arc (Step by Step)
- Muscle tension increases (due to active contraction or excessive passive stretch)
- GTO Ib afferent fibers fire, entering the spinal cord via the dorsal root
- Ib fibers synapse on Ib inhibitory interneurons in the spinal cord gray matter (first synapse)
- Ib inhibitory interneurons synapse on alpha motor neurons of the homonymous muscle (second synapse) - producing IPSPs (inhibitory postsynaptic potentials)
- Alpha motor neuron firing decreases
- Homonymous muscle relaxes (autogenic inhibition)
Simultaneously:
- Synergistic muscles also relax
- Antagonist muscles receive excitation (via excitatory interneurons) and contract - reciprocal excitation
This is the exact opposite pattern of the stretch reflex (which excites the homonymous muscle and inhibits the antagonist).
(Costanzo Physiology, 7th ed., pp. 111-112)
C. The Ib Inhibitory Interneuron - A Convergence Point
The Ib inhibitory interneuron is not exclusively driven by GTOs. It receives convergent input from:
- GTOs (primary input - Ib afferents)
- Muscle spindles (Ia and II afferents)
- Cutaneous receptors
- Joint receptors
- Descending pathways (both excitatory and inhibitory)
This convergence allows for sophisticated spinal control of muscle force. For example, during grasping a delicate object, cutaneous input from the fingertips reaching the surface can facilitate Ib interneuron activity, reducing muscle contractile force to allow a gentle grip.
(Kandel, Principles of Neural Science, 6th ed., p. 818)
D. Widespread Spinal Connections
The Ib fiber connections are not limited to the homonymous muscle. Ib fibers form widespread connections with motor neurons innervating muscles acting at different joints, making the GTO-Ib interneuron circuit part of broader spinal networks that coordinate multi-joint limb movements.
(Kandel, Principles of Neural Science, 6th ed., p. 818)
10. Functional Roles of the Golgi Tendon Reflex
A. Protective Function - The Lengthening Reaction
When tension on a muscle becomes extreme, the GTO-mediated inhibition becomes so powerful that it produces sudden, complete relaxation of the entire muscle - the lengthening reaction (also called the clasp-knife response in pathological states).
This likely evolved as a protective mechanism to prevent:
- Tearing of the muscle fibers
- Avulsion of the tendon from its bony attachment
(Guyton & Hall, p. 681)
B. Clasp-Knife Reflex (Pathological Exaggeration)
An exaggerated form of the Golgi tendon reflex seen in spasticity (upper motor neuron lesions with increased muscle tone):
- When a joint is passively flexed, the opposing hyperactive muscles initially resist strongly (high resistance phase)
- As passive flexion continues, tension builds until the GTO threshold is reached
- The Golgi tendon reflex fires, causing sudden relaxation - the joint then closes rapidly (release phase)
- This resembles a pocket knife closing: slow high resistance followed by sudden snap shut
(Costanzo Physiology, 7th ed., p. 112)
C. Equalization of Contractile Forces Among Motor Units
The GTO reflex helps distribute contractile activity evenly across muscle fibers within a muscle:
- Motor units generating excessive tension become inhibited via GTO feedback
- Less-loaded motor units take on a proportionally greater share
- This prevents over-fatigue or injury to any single subset of motor units
- Results in more uniform recruitment and load sharing
(Guyton & Hall, p. 682)
D. Precise Force Regulation During Normal Movement
Studies using population recordings from Ib axons in cat soleus during walking show that average GTO population discharge rate is a close index of total muscle force. The linear relationship between GTO firing frequency and muscle force confirms the GTO serves as a continuous, real-time force transducer, providing the CNS with accurate efference copy-independent force feedback for fine motor control.
(Kandel, Principles of Neural Science, 6th ed., p. 818)
11. Higher-Level Integration: Spindle and GTO Signals to the Brain
Signals from both the muscle spindle (Ia and II) and GTO (Ib) are transmitted to higher centers:
- Local spinal cord - for immediate reflex responses (as described above)
- Cerebellum via the spinocerebellar tracts - for coordination, timing, and smooth movement
- Cerebral cortex - via thalamocortical projections - contributing to conscious proprioception and fine voluntary motor control
The cerebellum uses spindle and GTO signals to:
- Compare intended vs. actual movement
- Issue corrective commands to spinal and cortical motor circuits
- Produce the timing and coordination of complex motor sequences
(Guyton & Hall, p. 682)
PART III: COMPARATIVE SUMMARY TABLE
| Feature | Muscle Spindle | Golgi Tendon Organ |
|---|---|---|
| Location | Within the muscle belly (in parallel) | Musculotendinous junction (in series) |
| Receptor type | Stretch receptor / length detector | Tension/force detector |
| Intrafusal fibers | Nuclear bag + nuclear chain | None (collagen fascicles + Ib nerve endings) |
| Primary afferent | Ia (17 µm) - annulospiral ending | Ib (16 µm) |
| Secondary afferent | II (8 µm) - flower spray ending | None |
| Motor innervation | Gamma (dynamic-d and static-s) | None |
| Detects | Muscle length and velocity of stretch | Muscle tension (force) |
| Best activated by | Passive stretch of muscle | Active muscle contraction |
| Dynamic response | Strong (especially from nuclear bag/Ia) | Present |
| Static response | Present (especially nuclear chain/II) | Present (proportional to tension) |
| Spinal reflex | Stretch reflex (monosynaptic, via Ia) | Golgi tendon reflex (disynaptic, via Ib interneuron) |
| Effect on homonymous muscle | Excitation (contraction) | Inhibition (relaxation) |
| Effect on antagonist | Inhibition (reciprocal inhibition) | Excitation (reciprocal excitation) |
| Synapse count | Monosynaptic (Ia-alpha) + polysynaptic (II) | Disynaptic (Ib-interneuron-alpha) |
| Clinical correlate | Knee jerk, biceps reflex, etc. | Clasp-knife reflex in spasticity |
| Protective role | Damps jerky contractions; maintains length | Prevents tendon avulsion; equalizes force |
| Higher-center projections | Cerebellum, cortex (proprioception) | Cerebellum, cortex (force feedback) |
PART IV: KEY CLINICAL CORRELATIONS
| Situation | Mechanism |
|---|---|
| Patellar (knee jerk) reflex | Tap patellar tendon -> stretches quadriceps spindles -> Ia fires -> monosynaptic excitation of quadriceps alpha MNs -> leg extends |
| Hyperreflexia in UMN lesions | Loss of supraspinal inhibition of spinal motor circuits -> increased alpha MN excitability -> exaggerated stretch reflexes + clonus |
| Hyporeflexia/areflexia in LMN lesions | Damage to afferent (Ia) or efferent (alpha MN) limb of the reflex arc -> absent or reduced DTRs |
| Clasp-knife sign | Spasticity -> increased muscle tone -> as joint passively moved, tension builds -> GTO threshold reached -> sudden relaxation (lengthening reaction) |
| Ataxia in cerebellar disease | Loss of spindle/GTO signal processing in cerebellum -> inability to compare intended vs. actual movement -> dysmetria, intention tremor |
| Muscle spindle in proprioception | Combined Ia + II spindle signals via thalamocortical pathways contribute to conscious awareness of joint position and movement (kinesthesia) |
Sources
- Guyton AC, Hall JE. Textbook of Medical Physiology, 14th ed. (ISBN 9780443111013). Chapter 55: Motor Functions of the Spinal Cord; the Cord Reflexes. pp. 676-682.
- Kandel ER et al. Principles of Neural Science, 6th ed. (ISBN 9781259642241). Chapter 32: Spinal Cord I. pp. 816-819 (Golgi Tendon Organs, Box 32-4; Fusimotor system and voluntary activity).
- Costanzo LS. Physiology, 7th ed. (ISBN 9780323793339). Chapter 3: Neurophysiology. pp. 111-112 (Golgi Tendon Reflex; Flexor-Withdrawal Reflex).
This is a shared conversation. Sign in to Orris to start your own chat.