Role of SNARE in NM transmission
Reading File
Finding Sources
Finding Sources
Searching PubMed
SNARE proteins neuromuscular junction synaptic vesicle exocytosis
Reading File
Reading File
Reading File
Reading File
Finding Sources
Loading Image
I now have everything needed for a thorough, well-illustrated answer. Here it is:
Role of SNARE Proteins in Neuromuscular (Synaptic) Transmission
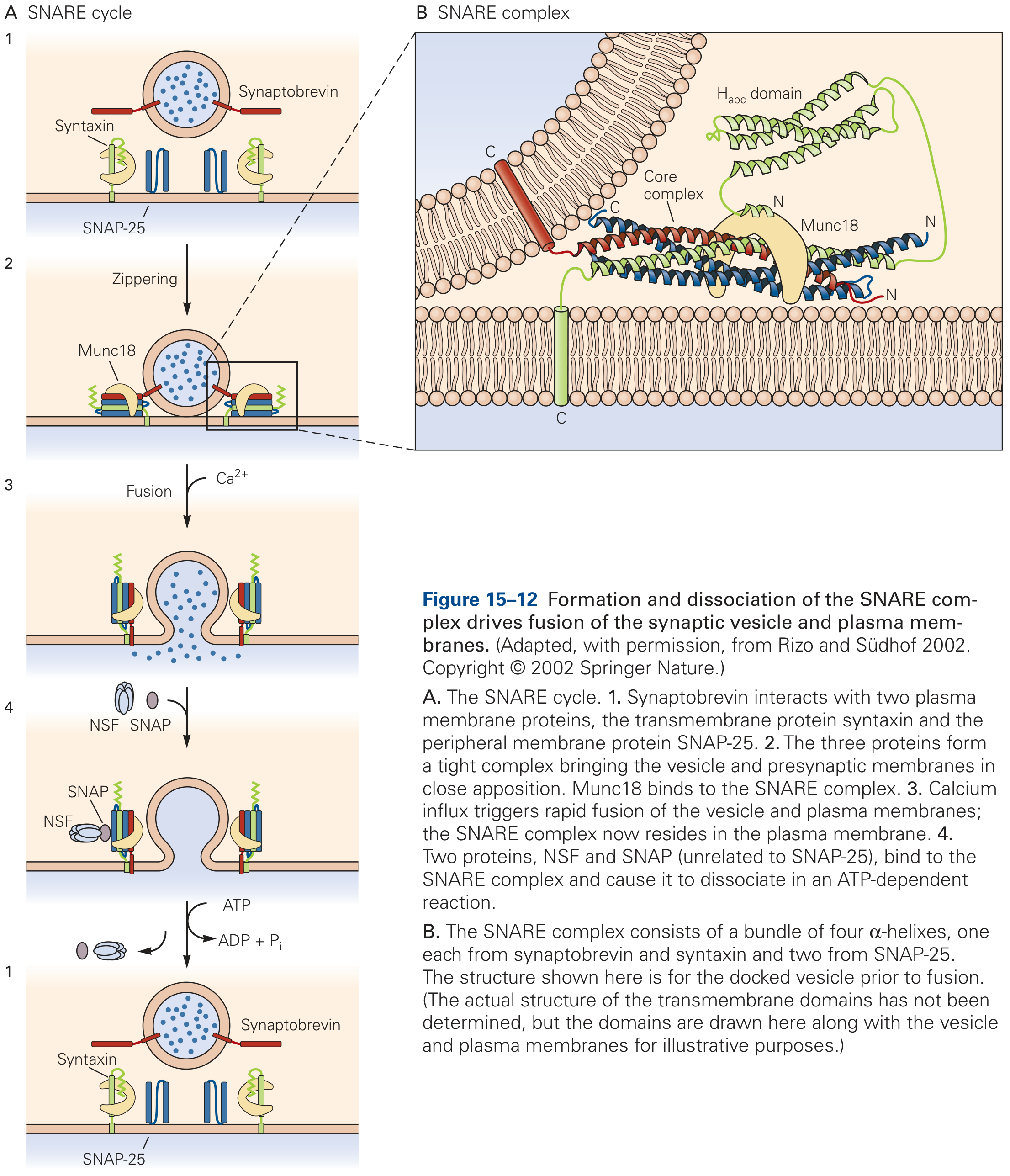
1. What Are SNAREs?
SNARE stands for Soluble N-ethylmaleimide-sensitive factor Attachment protein REceptors. They are a family of proteins universally involved in membrane fusion from yeast to humans. In the context of neuromuscular (and all synaptic) transmission, they are the core molecular machinery that drives acetylcholine (ACh) release from the motor nerve terminal.
SNAREs contain a conserved ~60-residue SNARE motif and are classified by location:
| Type | Also called | Location | Key residue |
|---|---|---|---|
| v-SNARE | R-SNARE | Synaptic vesicle | Arginine (R) |
| t-SNARE | Q-SNARE | Target (plasma) membrane | Glutamine (Q) |
2. The Three Key SNARE Proteins at the NMJ
At the neuromuscular junction and nerve terminals, three specific SNAREs drive vesicle fusion:
-
Synaptobrevin / VAMP (Vesicle-Associated Membrane Protein)
- v-SNARE; 18 kDa integral membrane protein on the synaptic vesicle
- Contributes one SNARE helix to the complex
-
Syntaxin-1
- t-SNARE; 33 kDa integral membrane protein anchored in the presynaptic plasma membrane
- Contributes one SNARE helix
-
SNAP-25 (Synaptosomal-Associated Protein, 25 kDa)
- t-SNARE; peripheral membrane protein tethered to the inner leaflet of the presynaptic membrane via palmitoylation
- Contributes two SNARE helices
Together, these four helices (1 from synaptobrevin + 1 from syntaxin + 2 from SNAP-25) zip into a tight 4-helix coiled-coil bundle called the trans-SNARE complex.
3. The SNARE Cycle - Step by Step
Step 1: Priming and Docking
- Synaptobrevin on the vesicle begins to interact with syntaxin and SNAP-25 on the plasma membrane at the active zone
- Munc18 (an SM protein - Sec1/Munc18-like) binds syntaxin before the SNARE complex assembles, acting as a chaperone; deletion of Munc18 abolishes all synaptic fusion
Step 2: SNARE Zippering
- The four SNARE motifs "zip" together from their N-termini toward their C-termini, like a zipper closing
- This zippering draws the vesicle and plasma membranes into close apposition, generating mechanical force that overcomes the activation energy barrier of membrane fusion
- The resulting ternary complex is extraordinarily stable
- The complex produces a prefusion intermediate and initiates the formation of a fusion pore (which flickers open and closed in electrophysiological recordings)
Step 3: Calcium Trigger via Synaptotagmin
- The SNARE complex alone is not sufficient for the rapid, Ca²⁺-regulated exocytosis at the NMJ
- When an action potential arrives and depolarizes the terminal, voltage-gated Ca²⁺ channels open
- Ca²⁺ binds synaptotagmin (a Ca²⁺ sensor on the vesicle) which then interacts with the assembled SNARE complex
- This triggers near-instantaneous, complete membrane fusion and ACh release into the synaptic cleft
Step 4: Complexin - The Safety Catch
- A small soluble protein called complexin binds the assembled SNARE complex
- It suppresses spontaneous (non-Ca²⁺) release while simultaneously enhancing Ca²⁺-evoked release - acting like a loaded spring
Step 5: Disassembly and Recycling (cis-SNARE to free SNARE)
- After fusion, the SNARE proteins are now in the same (plasma) membrane as a cis-SNARE complex
- The cytoplasmic ATPase NSF (N-ethylmaleimide-Sensitive Fusion protein) binds via the adaptor α-SNAP (Soluble NSF Attachment Protein; unrelated to SNAP-25)
- NSF uses ATP hydrolysis to disassemble the cis-SNARE complex, regenerating free SNAREs for the next round of exocytosis
4. Clinical Relevance: Botulinum and Tetanus Toxins
The SNARE proteins are the molecular targets of two of the most potent biological toxins known, which is strong evidence for their essential role:
| Toxin | Type | SNARE Target | Effect |
|---|---|---|---|
| Botulinum toxin (BoNT) B, D, F, G | Zn²⁺-protease | Synaptobrevin | Cleaves v-SNARE |
| Botulinum toxin A, C, E | Zn²⁺-protease | SNAP-25 | Cleaves t-SNARE |
| Botulinum toxin C | Zn²⁺-protease | Syntaxin | Cleaves t-SNARE |
| Tetanus toxin | Zn²⁺-protease | Synaptobrevin | Cleaves v-SNARE (in inhibitory neurons) |
Cleavage of any one SNARE protein prevents the trans-SNARE complex from forming, blocking ACh release at the NMJ and causing flaccid paralysis (botulism) or spastic paralysis (tetanus, through blocking inhibitory interneurons).
Therapeutic applications of BoNT-A/B: Injected locally to treat blepharospasm, strabismus, cervical dystonia, hyperhidrosis, chronic migraine, and spasticity.
5. Summary
Action potential → Ca²⁺ influx → Ca²⁺ binds synaptotagmin
↓
Synaptotagmin activates SNARE complex (synaptobrevin + syntaxin + SNAP-25)
↓
4-helix coiled-coil zippering → membranes fused → fusion pore opens
↓
ACh released into synaptic cleft → muscle depolarization
↓
NSF + α-SNAP + ATP → SNARE complex disassembled → recycled
The SNARE machinery is the universal and indispensable engine of regulated exocytosis at the neuromuscular junction.
Sources:
- Kandel's Principles of Neural Science, 6th Ed., pp. 392-394 (Chapter 15 - SNARE Proteins Catalyze Fusion)
- Histology: A Text and Atlas, 8th Ed., pp. 152-153 (Exocytosis and trans-SNARE complex)
- Goodman & Gilman's Pharmacological Basis of Therapeutics, p. 201 (Junctional Transmission - Molecular Mechanism of Exocytosis)
This is a shared conversation. Sign in to Orris to start your own chat.