Resting membrane potential
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
I now have excellent material from multiple authoritative textbooks. Here is a thorough answer:
Resting Membrane Potential
Definition
The resting membrane potential (RMP) is the electrical potential difference across the membrane of an excitable cell (nerve, muscle) when it is not generating a signal. By convention, the intracellular potential is expressed relative to the extracellular fluid (which is set to 0 mV). The inside of the cell is negative relative to the outside.
Typical values:
- Neurons: -65 to -70 mV (commonly cited as -70 mV)
- Skeletal/cardiac muscle fibers: -70 to -90 mV
This is what it looks like when measured with a microelectrode:
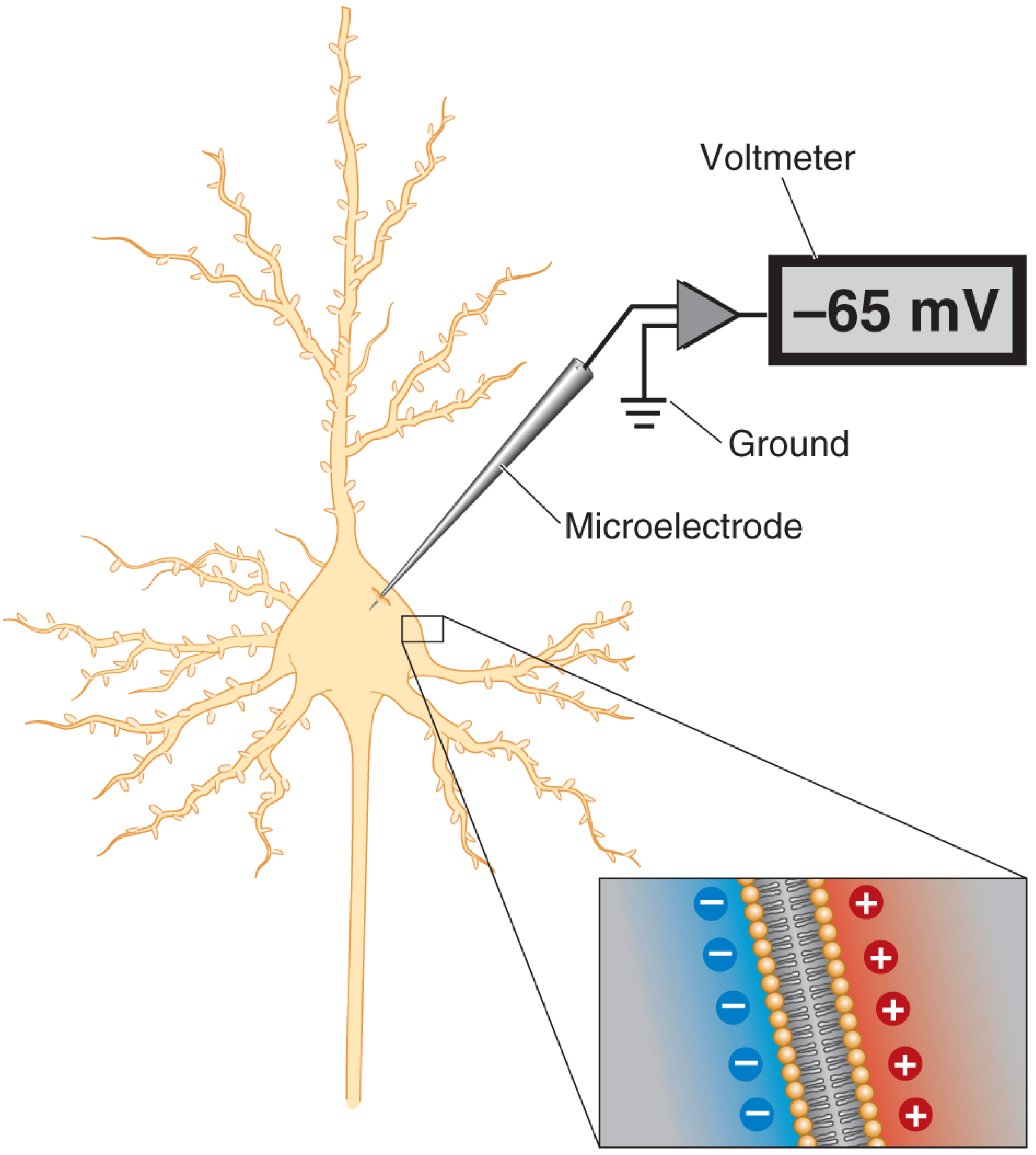
Measuring the resting membrane potential. A voltmeter measures the difference in electrical potential between the tip of a microelectrode inside the cell and a reference wire in extracellular fluid. The inside is about -65 mV relative to outside. - Neuroscience: Exploring the Brain, 5th Ed.
Ionic Basis: The Three Pillars
1. Ion Concentration Gradients (set by the Na⁺-K⁺ ATPase)
The Na⁺-K⁺ pump continuously maintains steep concentration gradients:
| Ion | Intracellular | Extracellular |
|---|---|---|
| Na⁺ | 14 mEq/L | 142 mEq/L |
| K⁺ | 140 mEq/L | 4 mEq/L |
These gradients mean K⁺ wants to leave the cell (high inside → low outside) and Na⁺ wants to enter (high outside → low inside). - Guyton & Hall Medical Physiology
2. Differential Membrane Permeability
At rest, the membrane is far more permeable to K⁺ and Cl⁻ than to Na⁺ and Ca²⁺. This is the single most important factor.
- K⁺ "leak" channels (tandem pore domain / TREK channels) are open at rest
- The membrane is about 100 times more permeable to K⁺ than to Na⁺ at rest
Because K⁺ leaks out down its concentration gradient, positive charges are lost from inside, making the interior negative. This negative charge then creates an electrical force pulling K⁺ back in - an equilibrium is reached at the K⁺ equilibrium potential (calculated by the Nernst equation: approximately -94 mV).
The small inward Na⁺ leak (through those same K⁺ leak channels) partially offsets this, bringing the actual resting potential up to about -86 mV by diffusion alone (Goldman equation result). - Guyton & Hall Medical Physiology
3. The Na⁺-K⁺ ATPase (Electrogenic Contribution)
The pump has two roles:
| Role | Mechanism | Magnitude |
|---|---|---|
| Indirect (major) | Maintains K⁺ and Na⁺ gradients that power the diffusion potentials | Most of the -86 mV |
| Direct (minor) | Pumps 3 Na⁺ out for only 2 K⁺ in → net outward positive charge | ~-4 mV additional |
The net result: approximately -90 mV in a large nerve fiber.
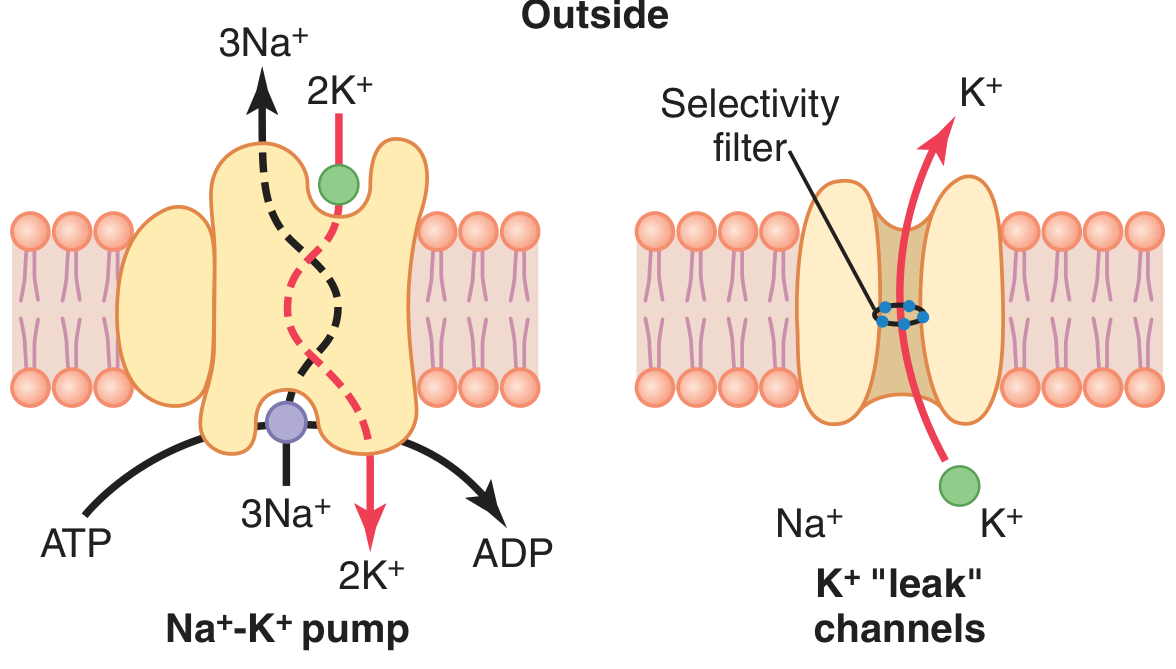
The Na⁺-K⁺ pump (left) and K⁺ "leak" channels (right). The pump's 3:2 stoichiometry makes it electrogenic. - Guyton & Hall Medical Physiology
Step-by-Step Build-Up (Guyton's Three Conditions)
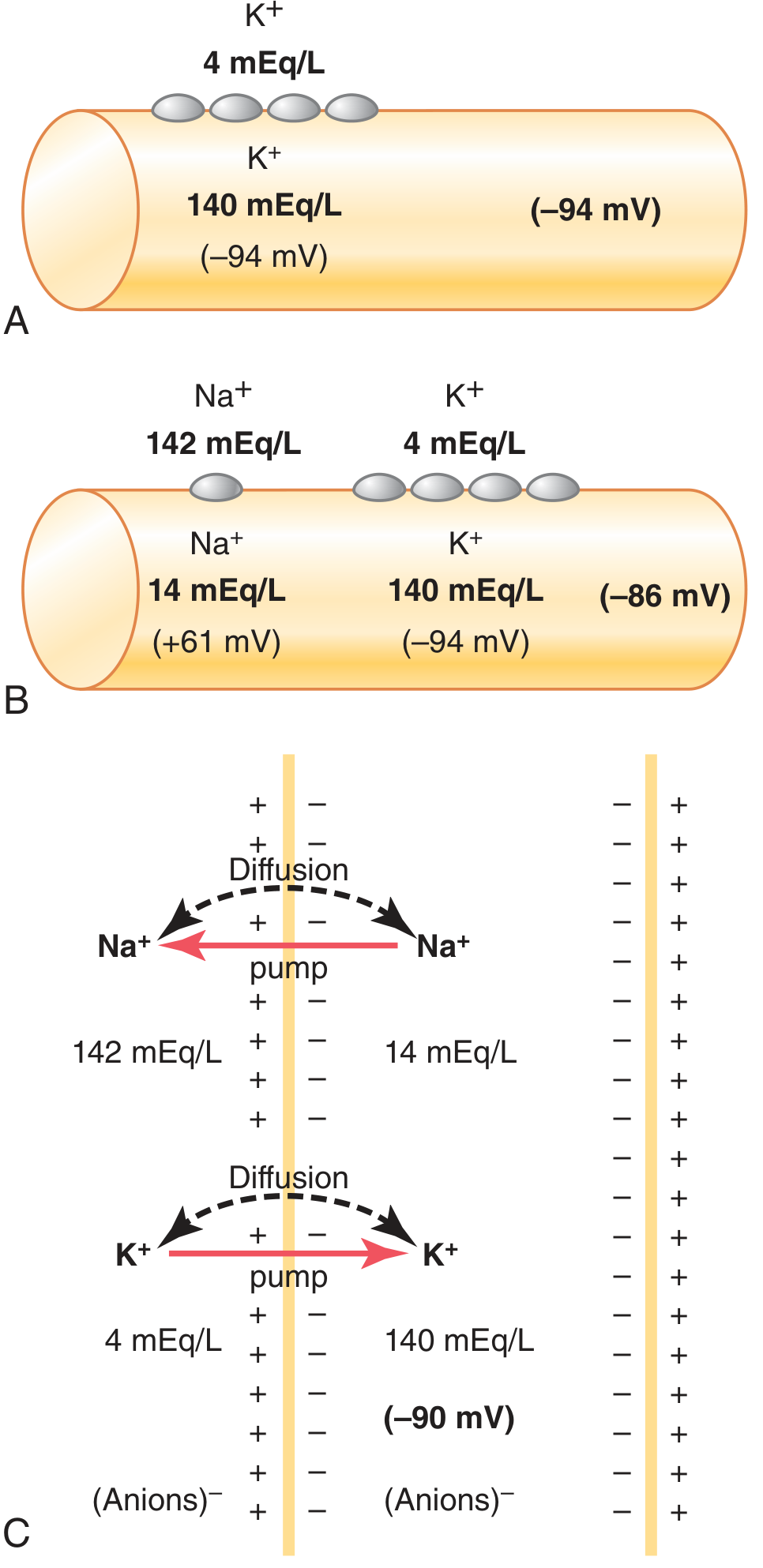
Guyton & Hall Medical Physiology - Establishment of resting membrane potential under three conditions.
- (A) K⁺ diffusion only: K⁺ inside/outside ratio = 35:1 → Nernst potential = -94 mV
- (B) K⁺ + Na⁺ diffusion: Na⁺ leak partially depolarizes → Goldman equation gives -86 mV
- (C) All factors including the Na⁺-K⁺ pump: Electrogenic pump adds ~-4 mV → net -90 mV
The Nernst and Goldman Equations
Nernst Equation - calculates the equilibrium potential for a single ion:
$$E_{ion} = \frac{61}{z} \log \frac{[ion]{outside}}{[ion]{inside}}$$
Where z = valence. For K⁺ (z = +1): E_K = 61 × log(4/140) ≈ -94 mV
For Na⁺ (z = +1): E_Na = 61 × log(142/14) ≈ +61 mV
Goldman (Constant Field) Equation - accounts for multiple ions and their relative permeabilities (P):
$$V_m = 61 \log \frac{P_{K^+}[K^+]o + P{Na^+}[Na^+]o + P{Cl^-}[Cl^-]i}{P{K^+}[K^+]i + P{Na^+}[Na^+]i + P{Cl^-}[Cl^-]_o}$$
Chord Conductance Equation - weights each ion's equilibrium potential by its relative conductance:
$$E_m = \frac{G_{K^+}}{G_T}E_{K^+} + \frac{G_{Na^+}}{G_T}E_{Na^+} + \frac{G_{Cl^-}}{G_T}E_{Cl^-} + \frac{G_{Ca^{2+}}}{G_T}E_{Ca^{2+}}$$
Ions with the highest conductance pull the membrane potential toward their own equilibrium potential. At rest, K⁺ conductance dominates, so RMP ≈ E_K. - Costanzo Physiology, 7th Ed.
Summary Table
| Factor | Direction of influence | Magnitude of contribution |
|---|---|---|
| K⁺ leak out (diffusion) | Hyperpolarizes (more negative) | Major (~-86 mV from diffusion) |
| Na⁺ leak in (diffusion) | Depolarizes (less negative) | Small offset |
| Cl⁻ distribution | Near equilibrium at RMP | Minor |
| Na⁺-K⁺ pump (electrogenic) | Hyperpolarizes | ~-4 mV |
| Intracellular anions (proteins) | Stabilize negativity inside | Passive contribution |
Clinical Relevance
- Hyperkalemia (high extracellular K⁺): reduces the K⁺ concentration gradient → RMP becomes less negative (depolarized) → cells become more excitable initially, then inexcitable (inactivation of Na⁺ channels). This explains muscle weakness and cardiac arrhythmias in hyperkalemia.
- Hypokalemia (low extracellular K⁺): increases the K⁺ gradient → RMP becomes more negative (hyperpolarized) → cells are harder to excite → muscle weakness, paralysis, arrhythmias.
- Local anesthetics block Na⁺ channels, preventing depolarization from reaching threshold from the RMP.
As Costanzo Physiology notes: the Na⁺-K⁺ ATPase is necessary both to create/maintain the K⁺ concentration gradient (and thus the RMP) AND to maintain the Na⁺ gradient (needed for the action potential upstroke).
Action potential in general physiology
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Finding Sources
Reading File
Reading File
I now have comprehensive material from Guyton, Costanzo, and supporting textbooks. Here is the full answer:
Action Potential - General Physiology
Definition
An action potential (AP) is a brief, rapid, self-propagating reversal of membrane potential that occurs in excitable cells (neurons, skeletal muscle, cardiac muscle). It is the fundamental electrical signal used by the nervous system to transmit information and by muscle to initiate contraction. - Costanzo Physiology, 7th Ed.
Starting point: RMP ≈ -70 mV (inside negative relative to outside).
Key Terminology
| Term | Definition |
|---|---|
| Depolarization | Membrane potential becomes less negative (inward positive current) |
| Hyperpolarization | Membrane potential becomes more negative (outward positive current) |
| Threshold potential | The critical voltage (~-55 mV) at which AP becomes inevitable; net inward current exceeds net outward current |
| Overshoot | Phase where the membrane potential becomes positive (cell interior briefly positive) |
| Undershoot (after-hyperpolarization) | Post-AP phase where Vm is more negative than RMP |
| Refractory period | Period during which another AP cannot be normally elicited |
Phases of the Action Potential
The AP of nerve and skeletal muscle proceeds through distinct, sequential phases:
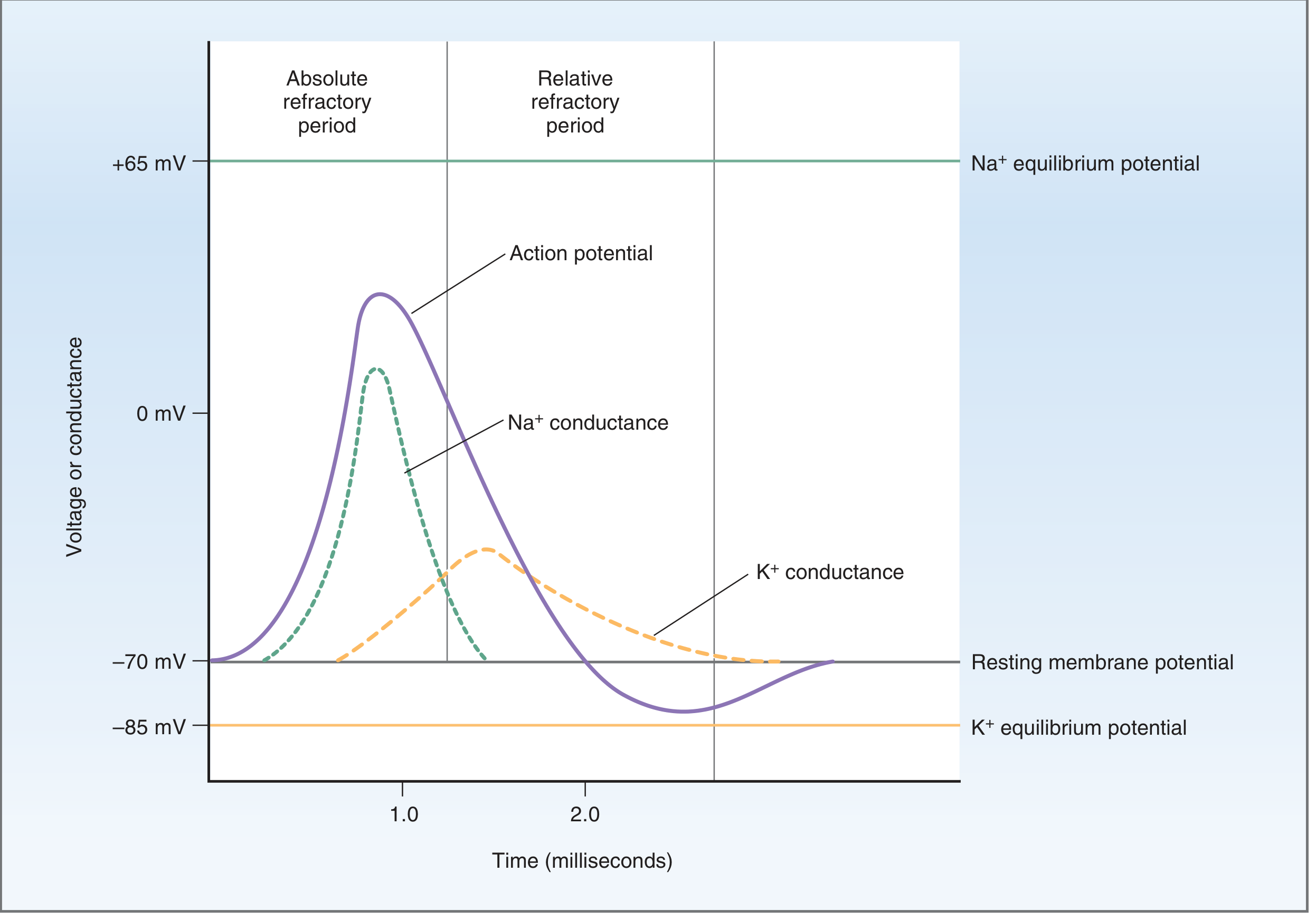
Time course of voltage and conductance changes during the action potential of nerve. - Costanzo Physiology, 7th Ed.
Phase 1 - Resting State (-70 mV)
- Membrane is polarized: inside negative, outside positive
- K⁺ leak channels open; membrane highly permeable to K⁺
- Voltage-gated Na⁺ channels are closed but available (activation gate closed, inactivation gate open)
- Voltage-gated K⁺ channels closed
Phase 2 - Upstroke (Depolarization) to threshold → ~+30 mV
- A stimulus depolarizes the membrane from -70 mV to threshold (~-55 mV, i.e., about 15-30 mV above RMP)
- At threshold, a positive feedback cycle is triggered: membrane depolarizes → voltage-gated Na⁺ channels open → Na⁺ rushes in → more depolarization → more Na⁺ channels open
- Na⁺ conductance increases up to 500-5000-fold
- Na⁺ floods in down its electrochemical gradient (E_Na = +61 mV), driving Vm toward E_Na
- In large fibers, Vm overshoots beyond 0 mV to approximately +30 to +40 mV
- The K⁺ conductance is still low at this point
Phase 3 - Repolarization
Two simultaneous events end the upstroke and restore the negative Vm:
- Na⁺ channel inactivation: The inactivation gate (slow gate) closes a few ten-thousandths of a second after opening. Na⁺ channels enter an inactivated (closed) state. Na⁺ conductance rapidly falls.
- Voltage-gated K⁺ channel activation: Delayed opening (slower than Na⁺ channels) increases K⁺ conductance ~30-fold. K⁺ rushes out of the cell, restoring negativity.
The combined effect: K⁺ conductance >> Na⁺ conductance → rapid repolarization back toward -70 mV.
Phase 4 - After-Hyperpolarization (Undershoot)
- Voltage-gated K⁺ channels remain open slightly longer than needed
- Vm is transiently driven below RMP, toward E_K (-85 to -94 mV)
- K⁺ channels gradually close → Vm returns to -70 mV (RMP)
The Voltage-Gated Na⁺ Channel - Three States
The Na⁺ channel has two gates: a fast activation gate (outer, opens quickly with depolarization) and a slow inactivation gate (inner, closes slowly with depolarization).
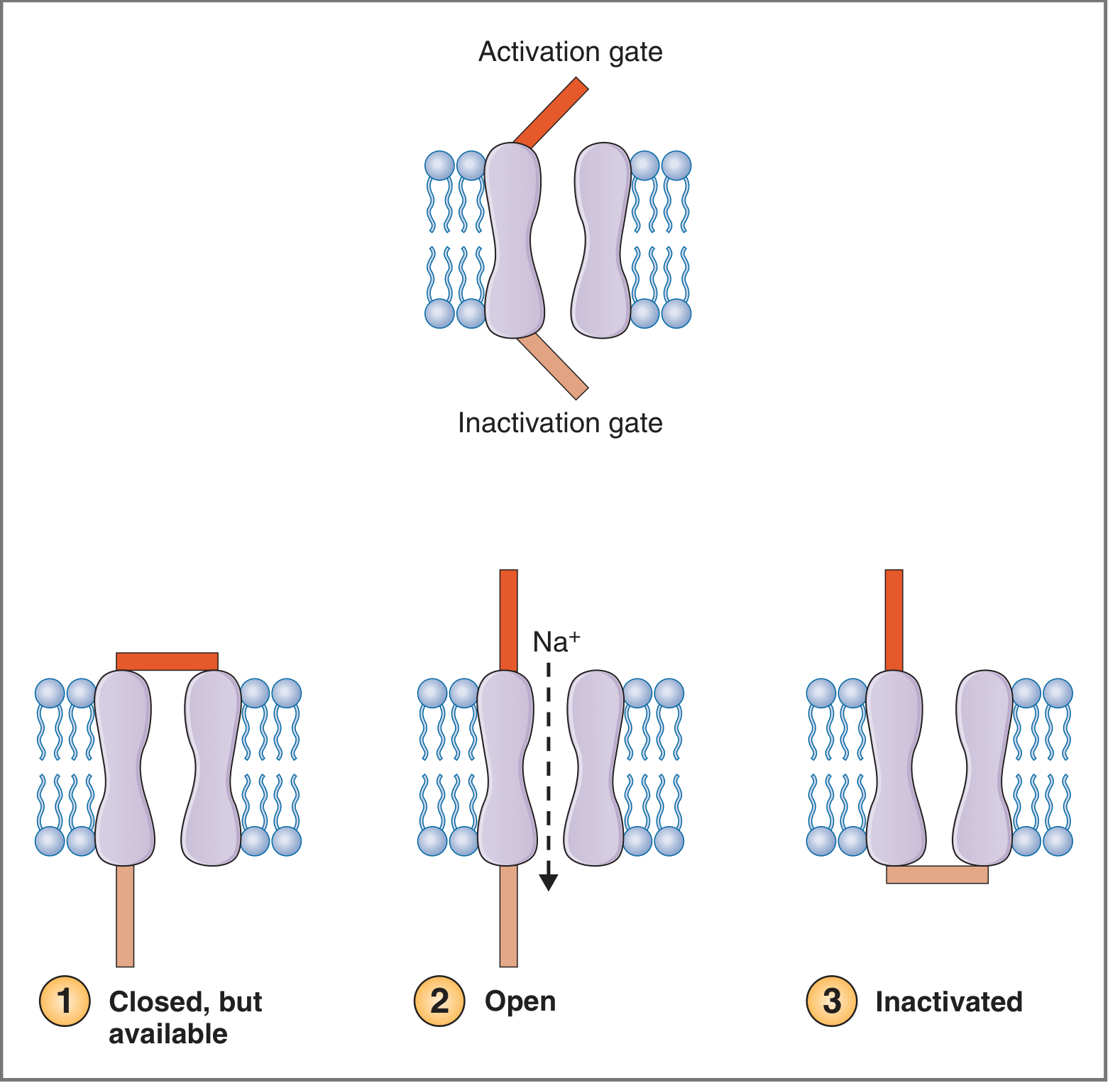
States of activation and inactivation gates on the nerve Na⁺ channel. - Costanzo Physiology, 7th Ed.
| State | Activation Gate | Inactivation Gate | Na⁺ flow | When |
|---|---|---|---|---|
| Closed, available | Closed | Open | No | At rest |
| Open | Open | Open | Yes (inward) | Upstroke |
| Inactivated | Open | Closed | No | Peak of AP |
- Recovery from inactivation requires repolarization back to near RMP - this is the basis of the absolute refractory period.
- Blocked by tetrodotoxin (TTX) from puffer fish and local anesthetics (lidocaine, bupivacaine). - Costanzo Physiology, 7th Ed.
Na⁺/K⁺ Conductance Changes - The Full Picture
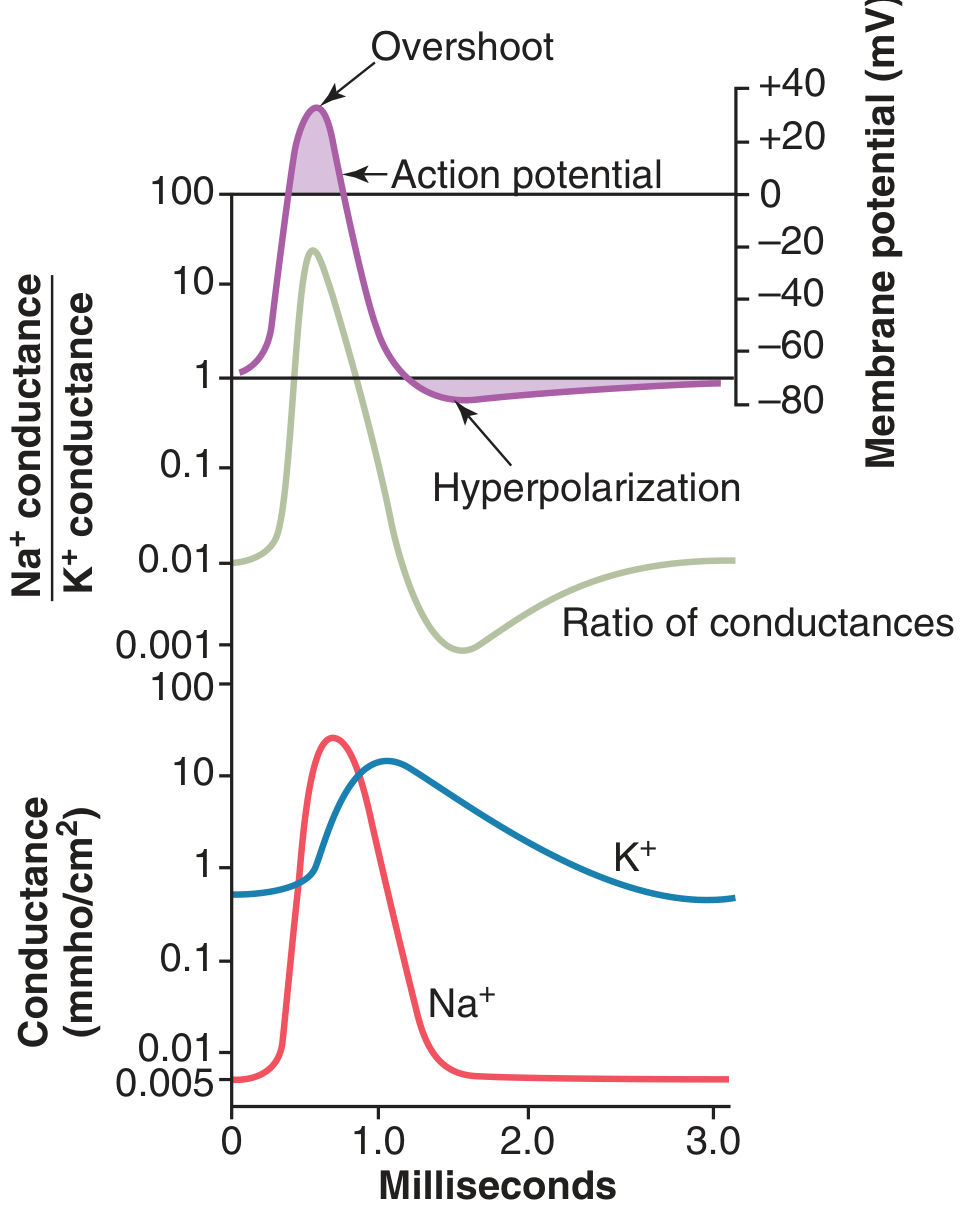
Changes in Na⁺ and K⁺ conductance during the action potential. Na⁺ conductance increases several thousand-fold during the early stages; K⁺ conductance increases ~30-fold during the latter stages. - Guyton & Hall Medical Physiology
Key sequence:
- Na⁺ conductance: spikes sharply at AP onset (>>K⁺), then rapidly falls as inactivation closes channels
- K⁺ conductance: rises slowly, peaks after Na⁺ channels are mostly inactivated, persists slightly into after-hyperpolarization
- Na⁺:K⁺ conductance ratio: >1000:1 at upstroke (driving depolarization) → reverses sharply during repolarization
Three Cardinal Characteristics of Action Potentials
1. Stereotypical Size and Shape
Each AP for a given cell looks identical: same peak voltage, same duration. The shape does not vary with stimulus strength.
2. All-or-None Response
Either the AP fires fully (threshold is reached) or it does not fire at all. A sub-threshold stimulus produces only a graded, decremental local potential that fades out. There is no "partial" AP.
3. Nondecremental Propagation
Each AP regenerates itself at each adjacent point on the membrane at full amplitude. It does not diminish as it travels. - Costanzo Physiology, 7th Ed.
Propagation of the Action Potential
When an AP fires at one point, local currents flow from the active (depolarized) site to adjacent resting membrane. These currents depolarize the adjacent segment to threshold, triggering a new AP there. This repeats continuously along the fiber.
Why does the AP travel in only one direction? The region behind the AP is in its absolute refractory period (Na⁺ channels inactivated), so it cannot fire again. Propagation therefore moves exclusively forward.
Conduction velocity is influenced by:
- Fiber diameter - larger fibers = faster conduction (less internal resistance)
- Myelination - saltatory conduction in myelinated fibers (AP jumps node to node) is far faster than continuous conduction in unmyelinated fibers
Refractory Periods
Absolute Refractory Period (ARP)
- Corresponds to the upstroke + most of repolarization
- No stimulus, however large, can trigger another AP
- Mechanism: Na⁺ channel inactivation gates are closed; channels cannot reopen until repolarization returns them to the "closed but available" state
- Sets the maximum firing frequency of a neuron
Relative Refractory Period (RRP)
- Follows ARP; coincides with the after-hyperpolarization
- An AP can be triggered, but requires a stronger-than-normal stimulus
- Mechanism: K⁺ conductance is still elevated; Vm is more negative than RMP; more inward current needed to reach threshold
Initiation of the AP - Positive Feedback Loop
The AP initiation is an explosive, self-sustaining positive feedback cycle:
Depolarization to threshold → Na⁺ channels open → Na⁺ influx → further depolarization → more Na⁺ channels open → ... (until all channels open)
This only becomes self-sustaining when Na⁺ influx > K⁺ efflux. Below threshold, K⁺ efflux wins and the membrane repolarizes without firing. - Guyton & Hall Medical Physiology
Role of Ca²⁺
- Ca²⁺ stabilizes Na⁺ channel gating by binding to the external surface of the channel protein
- Hypocalcemia removes this stabilizing effect → Na⁺ channels activate at lower voltages → membrane becomes hyperexcitable → spontaneous discharge → tetany
- Voltage-gated Ca²⁺ channels ("slow channels") also contribute to depolarization in some cells (e.g., sinoatrial node, smooth muscle) - Guyton & Hall Medical Physiology
Pharmacological Blockers (Summary)
| Drug/Toxin | Target | Effect |
|---|---|---|
| Tetrodotoxin (TTX) | Voltage-gated Na⁺ channels | Blocks upstroke; prevents AP |
| Local anesthetics (lidocaine, bupivacaine) | Voltage-gated Na⁺ channels | Blocks upstroke; nerve block |
| Tetraethylammonium (TEA) | Voltage-gated K⁺ channels | Blocks repolarization |
Quick Comparison: Nerve AP vs. Cardiac AP
| Feature | Nerve / Skeletal Muscle | Cardiac Ventricular |
|---|---|---|
| RMP | ~-70 mV | ~-90 mV |
| Duration | ~1-2 ms | ~200-400 ms |
| Plateau phase | Absent | Present (Phase 2, Ca²⁺ channels) |
| ARP | Short | Very long (prevents tetany) |
| Pacemaker | No | SA node (spontaneous) |
Ficks law of diffusion
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Loading Image
I now have thorough, multi-source material. Here is the complete answer:
Fick's Law of Diffusion
Background
Adolf Fick (1829-1901) was a German physiologist who in 1855 formulated the quantitative laws governing the passive movement of molecules down a concentration gradient. These laws are fundamental to physiology, pharmacology, and respiratory medicine.
Diffusion is the net movement of molecules from a region of high concentration to a region of low concentration, driven entirely by random thermal motion - no energy (ATP) is required.
Fick's First Law (Steady-State Diffusion)
Fick's First Law describes diffusion under steady-state conditions (when the concentration gradient is not changing over time):
$$\boxed{J = -D \cdot \frac{\Delta C}{\Delta x}}$$
Or in the physiological membrane form:
$$\boxed{\dot{V}_x = \frac{D \cdot A \cdot \Delta P}{\Delta x}}$$
| Symbol | Meaning | Units |
|---|---|---|
| J | Flux (net diffusion rate) | mol/cm²/s |
| D | Diffusion coefficient | cm²/s |
| ΔC | Concentration difference (C₁ - C₂) | mol/L |
| Δx | Thickness of the membrane/diffusion path | cm |
| A | Surface area available for diffusion | cm² |
| ΔP | Partial pressure difference (for gases) | mmHg |
| − | Negative sign indicates flow is down the gradient |
The negative sign indicates net flux occurs in the direction of decreasing concentration. Net flux is zero when ΔC = 0 (equilibrium). - Fitzpatrick's Dermatology
Fick's Second Law (Non-Steady-State Diffusion)
Fick's Second Law predicts flux when the concentration gradient is changing over time:
$$\frac{\partial C}{\partial t} = D \cdot \frac{\partial^2 C}{\partial x^2}$$
It also gives the relationship between time and distance of diffusion:
$$\Delta t = \frac{x^2}{2D}$$
This has a profound physiological implication: diffusion time scales with the square of distance.
| Distance | Time (water molecule, D = 2.5 × 10⁻⁵ cm²/s) |
|---|---|
| 10 μm (width of stratum corneum) | ~0.4 ms - very fast |
| 100 μm | ~40 ms - much slower |
| 1 mm | ~4 seconds |
| 1 cm | ~400 seconds |
This is why diffusion is effective across short distances (cell membranes, alveolar walls) but completely impractical over long distances - which is why the circulatory system exists. - Fitzpatrick's Dermatology
The Diffusion Coefficient (D)
D reflects how easily a molecule moves through a particular medium. It depends on:
| Factor | Effect on D | Rationale |
|---|---|---|
| Temperature ↑ | D ↑ | Greater thermal kinetic energy |
| Molecular weight ↑ | D ↓ | Larger/heavier molecules move slower (D ∝ 1/√MW) |
| Viscosity of medium ↑ | D ↓ | More resistance to movement |
| Lipid solubility ↑ | D ↑ (for membranes) | Easier entry into lipid bilayer |
For gases, D is also multiplied by solubility in the medium (not just size). This is critical:
- CO₂ diffusion coefficient across biological membranes is ~20 times higher than O₂
- Despite being a larger molecule, CO₂ is far more soluble in water/tissue
- For the same partial pressure gradient, CO₂ diffuses 20× faster than O₂ - Costanzo Physiology, 7th Ed.; Miller's Anesthesia, 10th Ed.
Full Physiological Form of Fick's Law
For membrane transport (as used clinically):
$$\dot{V}_x = \frac{D \cdot A \cdot \Delta P}{\Delta x}$$
Rate of diffusion is:
- Directly proportional to: diffusion coefficient (D), surface area (A), concentration/partial pressure gradient (ΔP or ΔC)
- Inversely proportional to: membrane thickness (Δx)
Application to Pulmonary Gas Exchange
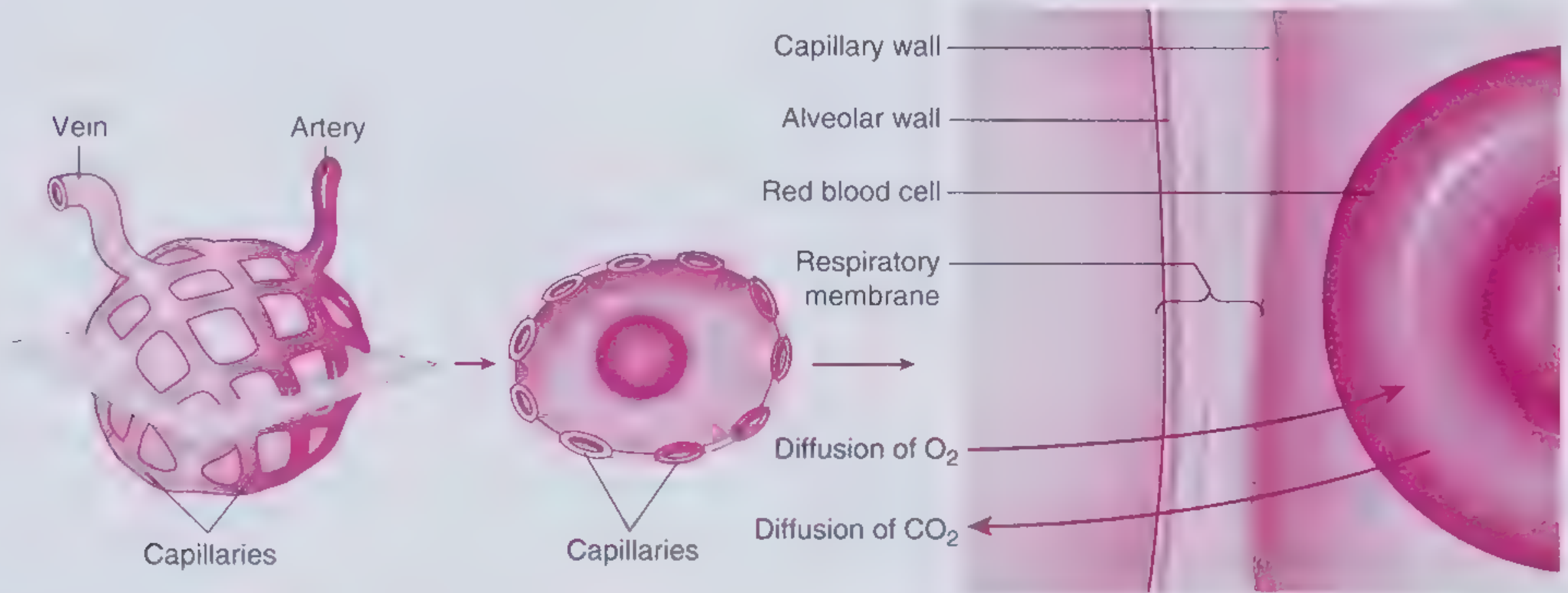
O₂ and CO₂ move across the alveolar-capillary membrane via Fick's laws of diffusion. - Frameworks for Internal Medicine
For O₂ across the alveolar-capillary membrane:
$$\dot{V}{O_2} = \frac{D \cdot A \cdot (P{A_{O_2}} - P_{c_{O_2}})}{\Delta x}$$
Where:
- P_AO₂ = alveolar PO₂ (~100 mmHg)
- P_cO₂ = pulmonary capillary PO₂ (~40 mmHg in mixed venous)
- Driving force = 60 mmHg for O₂
Lung Diffusing Capacity (D_L)
The terms D, A, and Δx can be combined into a single measurable parameter:
$$D_L = \frac{D \cdot A}{\Delta x}$$
So the equation simplifies to:
$$\dot{V}_x = D_L \cdot \Delta P$$
D_LCO (diffusing capacity for carbon monoxide) is the clinical measurement of lung diffusion capacity, because CO transfer is entirely diffusion-limited. - Costanzo Physiology, 7th Ed.
Factors That Change D_L (Clinical Relevance)
| Condition | Change in D_L | Mechanism |
|---|---|---|
| Emphysema | ↓ | Destruction of alveoli → ↓ surface area (A) |
| Pulmonary fibrosis | ↓ | Thickening of alveolar-capillary membrane → ↑ Δx |
| Pulmonary edema | ↓ | Increased diffusion distance → ↑ Δx |
| Anemia | ↓ | Less Hb to bind O₂ (reduces protein-binding component) |
| Exercise | ↑ | Recruitment of additional capillaries → ↑ A |
| Polycythemia | ↑ | More Hb available |
Why does impaired diffusion cause hypoxemia but NOT hypercapnia?
- CO₂ is ~20× more soluble in water than O₂ → its effective diffusion coefficient is far higher
- Even a thickened membrane transfers CO₂ adequately
- Hypoxemia often triggers hyperventilation, which further lowers PaCO₂
- Frameworks for Internal Medicine
Application to Drug Pharmacokinetics (Katzung)
Fick's Law in pharmacology:
$$\text{Flux} = \frac{\text{Area} \times \text{Permeability coefficient} \times (C_1 - C_2)}{\text{Thickness}}$$
- Lipid diffusion is the most important rate-limiting step for most drugs
- The lipid:aqueous partition coefficient determines how readily a drug enters lipid membranes
- Ionized drugs cannot diffuse well (charged molecules attract water, are lipid-insoluble)
- Henderson-Hasselbalch equation predicts the ionized fraction and therefore the diffusibility of weak acids/bases - Katzung's Basic & Clinical Pharmacology, 16th Ed.
Types of Diffusion in Physiology
| Type | Mechanism | Examples |
|---|---|---|
| Simple (passive) diffusion | Down concentration gradient, no carrier, no energy | O₂, CO₂, lipid-soluble drugs, ethanol |
| Facilitated diffusion | Carrier-mediated, down gradient, no energy | Glucose (GLUT transporters), amino acids |
| Aqueous diffusion | Through pores/channels, down gradient | Water (aquaporins), small ions |
| Active transport | Against gradient, requires ATP | Na⁺-K⁺ ATPase, drug efflux pumps (P-gp) |
Only simple/facilitated diffusion and aqueous diffusion follow Fick's law directly. Active transport does not, as it moves molecules against their concentration gradient.
Summary
$$\boxed{\text{Rate of diffusion} \propto \frac{D \times A \times \Delta C}{\Delta x}}$$
The key relationships to remember:
- More surface area → faster diffusion (lungs have ~70 m² for this reason)
- Thicker membrane → slower diffusion (fibrosis impairs gas exchange)
- Larger gradient → faster diffusion (breathing high-FiO₂ increases PAO₂ and thus the gradient)
- Higher D → faster diffusion (CO₂ >> O₂ per unit gradient)
- Diffusion is only efficient over micrometers - the capillary system brings blood to within 20-50 μm of every metabolizing cell
This is a shared conversation. Sign in to Orris to start your own chat.