VISION - Comprehensive Study Notes
Sources: Costanzo Physiology 7e; Ganong's Review of Medical Physiology 26e
PART I: Structures of the Eye and Their Functions
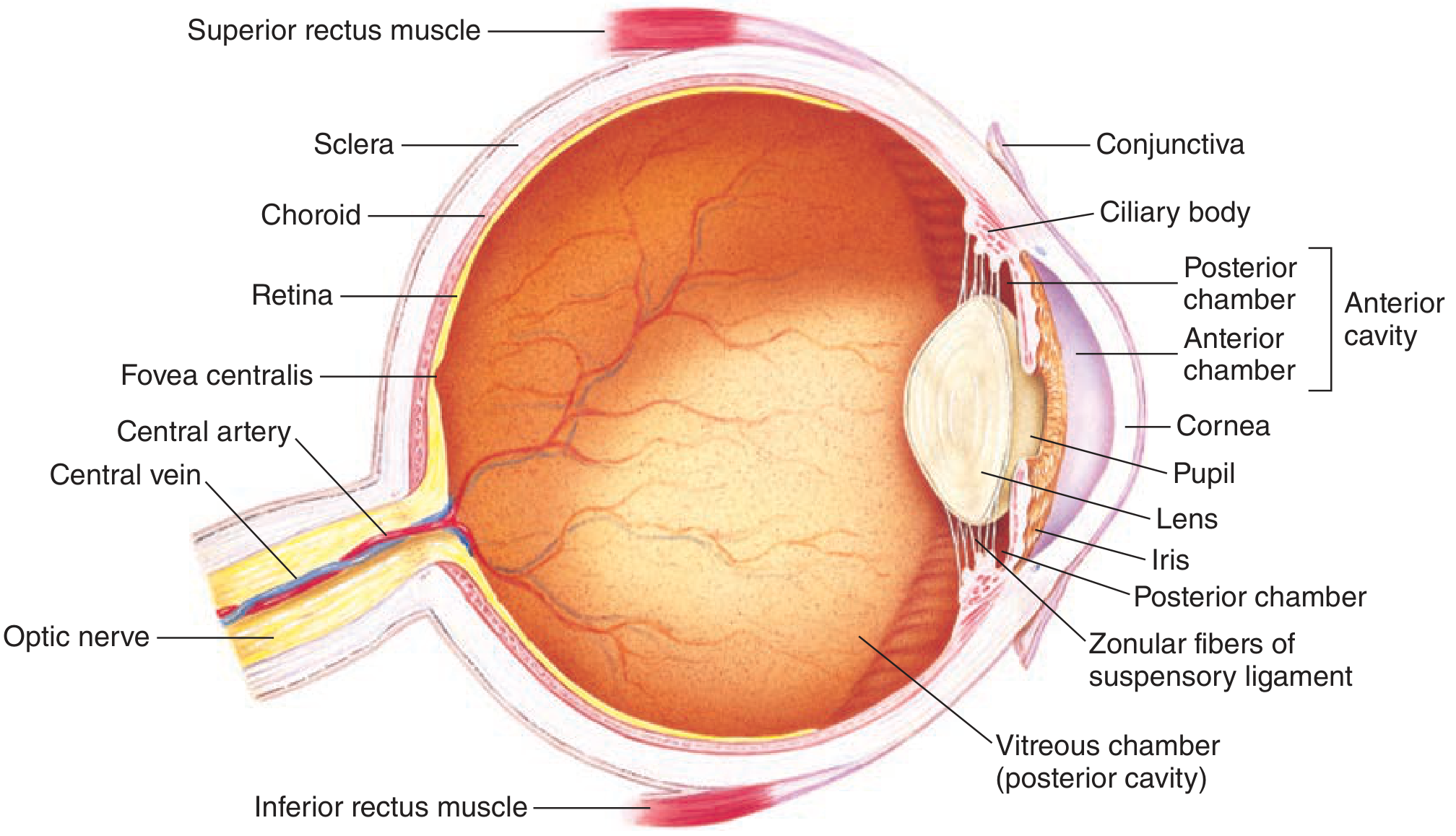
The wall of the eye is organized as three concentric layers:
Outer (Fibrous) Layer
| Structure | Function |
|---|
| Sclera | White, opaque, tough protective covering; no light passes through it |
| Cornea | Transparent anterior modification of the sclera; admits light and provides ~70% of total refractive power (~43 diopters); covered by the corneal epithelium |
| Conjunctiva | Clear mucous membrane covering the sclera; keeps the cornea moist |
Middle (Vascular) Layer - the Uvea
| Structure | Function |
|---|
| Choroid | Highly vascular; delivers oxygen and nutrients to eye structures, especially the outer retina |
| Ciliary body | Contains circular and longitudinal smooth muscle; controls lens shape for accommodation; produces aqueous humor |
| Iris | Pigmented, opaque diaphragm; contains sphincter muscle (parasympathetic - miosis) and radial dilator muscle (sympathetic - mydriasis); controls pupil diameter and thus retinal illumination (up to 16-fold change in light reaching the retina) |
| Pupil | The aperture in the iris |
Inner (Neural) Layer
| Structure | Function |
|---|
| Retina | Neural tissue containing photoreceptors (rods and cones); lines posterior 2/3 of the choroid; the site of phototransduction |
| Macula | Central retinal zone with highest visual acuity |
| Fovea | Depression in the macula where light is focused; contains only cones; highest acuity |
| Optic disc (blind spot) | Point where ganglion cell axons leave as the optic nerve; no photoreceptors, hence a blind spot |
Other Key Structures
| Structure | Function |
|---|
| Lens | Biconvex, transparent; focuses light; curvature adjusted by ciliary muscle via zonular fibers |
| Zonular fibers (suspensory ligament) | Connect lens to ciliary body; transmit ciliary muscle tension to the lens |
| Aqueous humor | Clear, protein-free fluid filling the anterior chamber (between cornea and lens); nourishes cornea and iris; produced by ciliary body, drains via canal of Schlemm |
| Vitreous humor | Gelatinous fluid filling the vitreous (posterior) chamber (between lens and retina); maintains eye shape |
PART I: Rods vs. Cones
| Feature | Rods | Cones |
|---|
| Light threshold | Low - extremely sensitive | High - require bright light |
| Vision type | Scotopic (night vision) | Photopic (day vision) |
| Acuity | Low | High |
| Color vision | No | Yes (three cone types: L, M, S) |
| Dark adaptation | Adapt late (slower) | Adapt early (faster) |
| Location | Peripheral retina (none in fovea) | Concentrated in fovea |
| Convergence | Many rods → one bipolar (high sensitivity, low acuity) | Few cones → one bipolar → one ganglion (low sensitivity, high acuity) |
| Photopigment amount | Large (free-floating discs) | Smaller (membrane infoldings) |
A single photon can activate a rod; several hundred photons are needed to activate a cone.
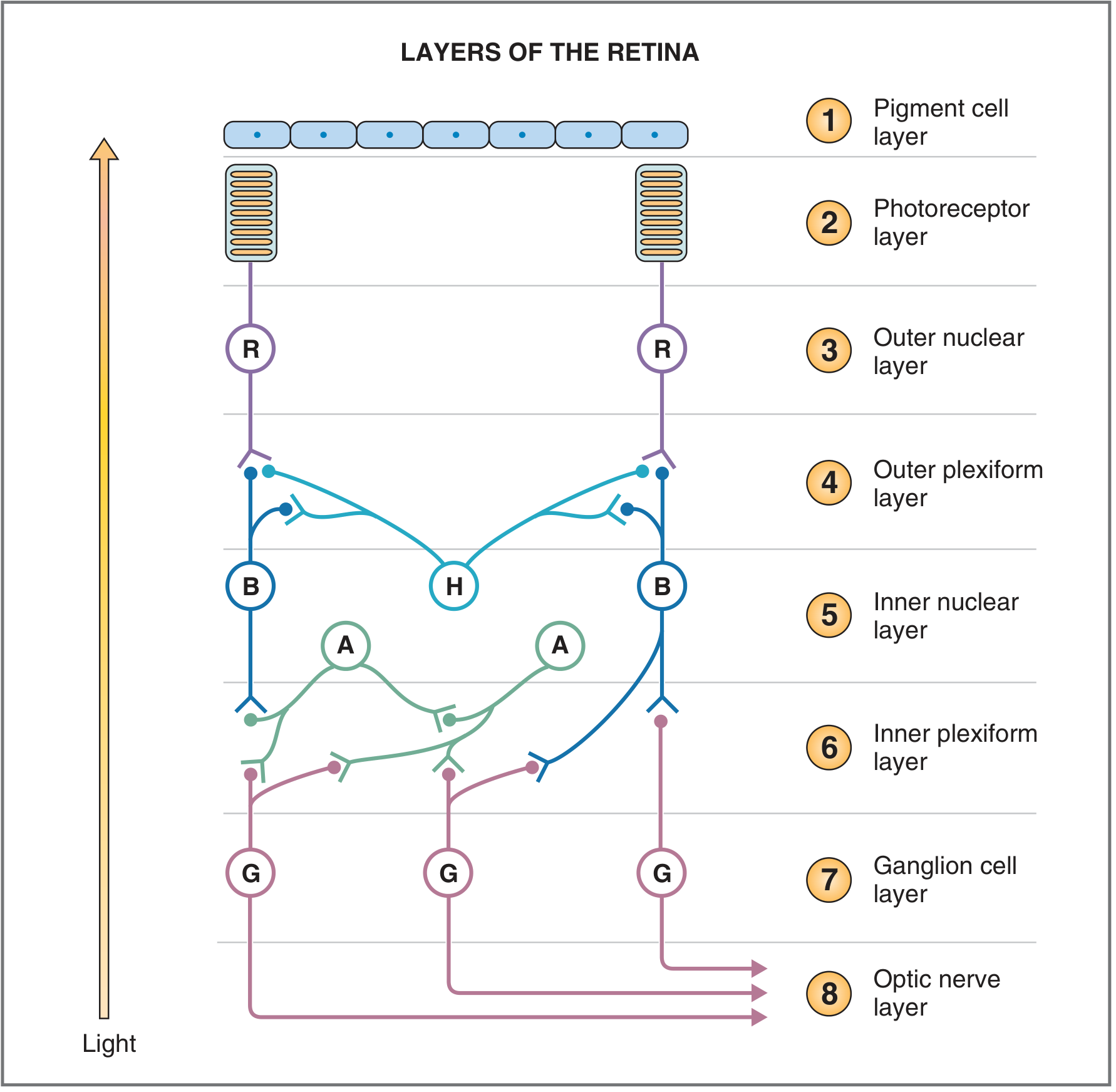
PART I: Layers of the Retina
Note: light enters from the bottom (vitreous side) and must pass through the ganglion and bipolar cell layers before reaching the photoreceptors at the back (pigment epithelium side).
- Pigment cell layer - absorbs stray light; converts all-trans retinal back to 11-cis retinal; extends tentacle-like processes into photoreceptors to prevent scatter
- Photoreceptor layer - outer and inner segments of rods and cones
- Outer nuclear layer - nuclei of photoreceptors
- Outer plexiform layer - first synaptic layer; photoreceptors → bipolar cells and horizontal cells
- Inner nuclear layer - cell bodies of bipolar (B), horizontal (H), and amacrine (A) cells
- Inner plexiform layer - second synaptic layer; bipolar and amacrine cells → ganglion cells
- Ganglion cell layer - cell bodies of retinal ganglion cells (the output neurons)
- Optic nerve layer - axons of ganglion cells forming the optic nerve
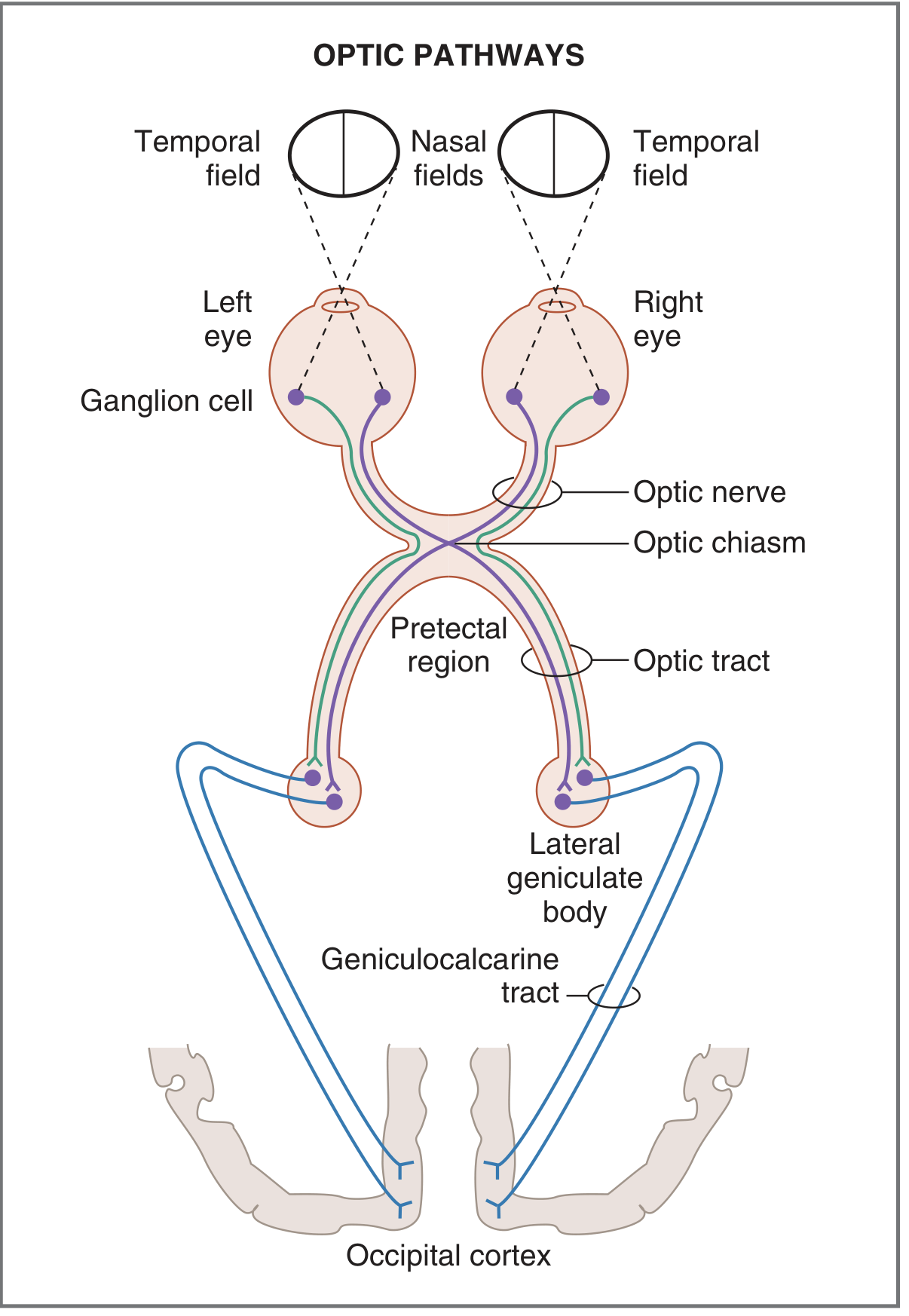
PART I: Neural Pathway - Rods and Cones to Visual Cortex
Step-by-step pathway:
- Photoreceptors (rods and cones) in the retina transduce light
- Bipolar cells - first-order neurons; relay signals from photoreceptors to ganglion cells via the outer plexiform layer
- Retinal ganglion cells - generate action potentials; their axons exit the eye at the optic disc and form the optic nerve (CN II)
- Optic chiasm - partial decussation: fibers from the nasal hemiretina of each eye cross to the opposite side; fibers from the temporal hemiretina stay ipsilateral
- Result: the left optic tract carries information from the right visual field (left temporal retina + right nasal retina)
- The right optic tract carries information from the left visual field
- Optic tract → Lateral geniculate nucleus (LGN) of the thalamus - relay nucleus; retains on-center/off-center patterns from ganglion cells
- Geniculocalcarine tract (optic radiation) - LGN neurons project to the primary visual cortex (V1)
- Primary visual cortex (V1) - located on both sides of the calcarine fissure in the occipital lobe
A small branch also projects to the pretectal region (pupillary light reflex) and to the superior colliculus (eye movement coordination).
PART I: Optics - How Light is Focused on the Retina
Light is refracted (bent) when passing between media of different densities. Refraction is the mechanism by which the eye focuses images on the retina. Refractive power is measured in diopters (D) = 1/focal length in meters.
Main refracting surfaces:
- Anterior corneal surface - provides ~70% of total refractive power (~43 D); its power is fixed
- Anterior and posterior surfaces of the lens - provide the remaining ~30% (~20 D at rest); power is adjustable
Light from objects >6 m away arrives as parallel rays and focuses at the principal focus. Rays from closer objects diverge and need more refractive power to focus on the retina.
Accommodation
Accommodation is the process by which the lens increases its curvature to increase refractive power, allowing the eye to focus on near objects.
Mechanism:
- Near object is viewed → diverging light rays require more power to focus
- Ciliary muscle contracts (parasympathetic, CN III)
- Contraction of the ciliary muscle releases tension on the zonular fibers
- The elastic lens, now released, becomes more spherical (rounder, increased curvature)
- Increased lens curvature = increased refractive power
- Near object is brought to sharp focus on the retina
Relaxed state (far vision): Ciliary muscle relaxed → zonular fibers taut → lens stretched flat → lower power → parallel rays from distant objects focused.
The accommodation response also includes convergence of the eyes and pupillary constriction (the accommodation triad/near triad).
PART I: Refractive Errors
| Condition | Problem | Correction |
|---|
| Myopia (nearsightedness) | Eyeball too long OR lens too curved → light focuses in front of retina; near objects seen clearly, distant objects blurred | Concave (diverging) lens |
| Hyperopia (farsightedness) | Eyeball too short OR lens too flat → light would focus behind retina; distant objects clearer but near objects blurred; requires constant accommodation even for far vision | Convex (converging) lens |
| Astigmatism | Cornea (or lens) has unequal curvature in different planes → different focal points; some parts of image in focus, others are not; causes blurring at all distances | Cylindrical lens oriented to match the abnormal curvature |
| Presbyopia | Loss of lens elasticity with aging → lens cannot become sufficiently spherical for accommodation → difficulty focusing on near objects; accommodation amplitude decreases progressively | Convex (reading) glasses; bifocals |
| Strabismus | Misalignment of the visual axes due to imbalance of extraocular muscle strength or innervation → eyes do not point at the same object simultaneously → diplopia or, in children, amblyopia (cortical suppression of one eye's image) | Glasses, patching, surgery |
PART II: Electrical Responses of Rods and Cones
Phototransduction - How Light is Converted to an Electrical Signal
Rhodopsin is the photopigment in both rods and cones, composed of:
- Opsin - a G protein-coupled receptor protein
- Retinal - aldehyde of vitamin A (chromophore)
Steps:
- Light strikes the outer segment → 11-cis retinal is photoisomerized to all-trans retinal (photoisomerization)
- Opsin undergoes conformational change → becomes activated (metarhodopsin II)
- Activated opsin activates transducin (a G protein, Gα subunit)
- Transducin activates phosphodiesterase (PDE)
- PDE cleaves cGMP → cGMP levels fall in the outer segment
- cGMP-gated cation channels close - in darkness these channels are held open by cGMP, allowing Na⁺ and Ca²⁺ inflow (the "dark current")
- Photoreceptor hyperpolarizes - closing channels stops inward current → membrane potential becomes more negative
- Less glutamate released from photoreceptor synaptic terminal (glutamate release is tonically high in the dark)
Key concept: Photoreceptors are depolarized in the dark and hyperpolarize in response to light. This is opposite to most sensory receptors.
Dark current: In darkness, cGMP-gated channels are open → continuous inflow of Na⁺ (and Ca²⁺) through the outer segment and outflow of K⁺ through the inner segment → maintained at approximately -40 mV.
Cone receptor potentials have a sharp onset and offset; rod receptor potentials have a sharp onset and slow offset. Rods are far more sensitive (respond at lower light levels).
The receptor potentials of photoreceptors (and most retinal cells) are graded local potentials, not action potentials. Only ganglion cells generate propagated all-or-none action potentials.
PART II: Electrical Responses of Bipolar, Horizontal, Amacrine, and Ganglion Cells
Bipolar Cells
Bipolar cells relay signals from photoreceptors to ganglion cells. They respond to glutamate released by photoreceptors and produce graded potentials only (no action potentials). There are two types:
- ON-bipolar cells - have metabotropic glutamate receptors (mGluR6); in the dark (high glutamate) they are hyperpolarized; when light hyperpolarizes the photoreceptor and reduces glutamate, the ON-bipolar cell depolarizes ("turned on" by light)
- OFF-bipolar cells - have ionotropic glutamate receptors (AMPA/kainate); in the dark (high glutamate) they depolarize; reduced glutamate with light causes hyperpolarization ("turned off" by light)
This creates the receptive field of bipolar cells with on-center/off-surround or off-center/on-surround patterns.
Horizontal Cells
- Located in the inner nuclear layer; processes run laterally in the outer plexiform layer
- Receive input from photoreceptors; provide lateral inhibitory feedback back to photoreceptors and to bipolar cells
- Respond to light with hyperpolarizing graded potentials
- Role: enhance contrast and sharpen spatial resolution; mediate the surround inhibition of bipolar cell receptive fields (when light hits the surround area, horizontal cells suppress the center response)
Amacrine Cells
- Located in the inner nuclear layer; processes run laterally in the inner plexiform layer
- Receive input from bipolar cells; modulate signaling between bipolar cells and ganglion cells
- Produce depolarizing potentials and action potential-like spikes (transient responses)
- Role: detect changes in luminance (motion detection); contribute to temporal processing; their mixed-polarity receptive fields (mixtures of on-center and off-center input) broaden the receptive fields of ganglion cells that receive primarily amacrine input
- Specific amacrine cell types: AII amacrine cells connect the rod pathway to cone bipolar cells in dim light conditions
Ganglion Cells
- The output neurons of the retina; the only retinal cells that fire propagated action potentials (all-or-none)
- Receive input from bipolar and amacrine cells
- Retain on-center/off-center receptive field organization when input is primarily from bipolar cells; acquire more diffuse receptive fields when input is primarily from amacrine cells
- Major classes:
- M (magnocellular / parasol) cells - large receptive fields; high contrast sensitivity; important for motion detection; project to magnocellular layers of LGN
- P (parvocellular / midget) cells - small receptive fields; high spatial acuity; color opponent; project to parvocellular layers of LGN
- Intrinsically photosensitive ganglion cells (ipRGCs) - contain melanopsin; project to suprachiasmatic nucleus (circadian rhythms) and pretectal area (pupillary reflex)
PART II: Visual Cortex - Cell Responses and Functional Organization
Primary Visual Cortex (V1 / Striate Cortex)
Located along the calcarine fissure in the occipital lobe. Like all neocortex it has six layers.
Inputs from LGN:
- Magnocellular pathway axons → terminate in layer 4Cα (then project to layer 4B)
- Parvocellular pathway axons → terminate in layer 4Cβ
- Interlaminar (koniocellular) pathway axons → terminate in layers 2 and 3 (blobs)
Cell types in V1:
| Cell Type | Response Properties |
|---|
| Layer 4 cells | On-center/off-surround or off-center/on-surround (like LGN cells); respond to spots of light; no orientation preference |
| Simple cells (layers 2, 3, 5, 6) | Respond to bars of light, lines, or edges of a specific orientation only; rotating the bar even 10° reduces firing; first level of feature detection |
| Complex cells | Prefer linear stimuli with a specific orientation; less dependent on exact position in visual field than simple cells; often respond maximally to a moving bar of the correct orientation |
| Hypercomplex cells | Respond best to lines of specific length, curves, or angles; end-stopped responses |
Organizational features of V1:
- Orientation columns - vertical columns (~1 mm diameter) where all cells prefer the same bar orientation; neighboring columns shift orientation preference by 5-10° per step through the full 360°
- Ocular dominance columns - alternate vertical stripes of cortex receiving input primarily from left vs. right eye; input from LGN is monocular in layer 4, but about half of cells in other layers are binocular
- Blobs - clusters of cells in layers 2 and 3 rich in cytochrome oxidase; dedicated to color processing; arranged in a mosaic pattern
Dorsal and Ventral Visual Pathways (Beyond V1)
Two major processing streams arise from V1:
| Pathway | Route | Function |
|---|
| Dorsal ("Where/How") pathway | V1 → V2 → V3 → MT (V5) → posterior parietal cortex (PP) | Spatial location, motion detection, visually guided action ("where is it?" and "how do I interact with it?") |
| Ventral ("What") pathway | V1 → V2 → V4 → inferotemporal cortex (IT) | Object recognition, shape, color, face recognition ("what is it?") |
The dorsal pathway uses mainly the magnocellular stream (motion, depth, fast processing). The ventral pathway uses mainly the parvocellular stream (color, fine detail, form). Color area V8 (part of V4) is specifically involved in color perception.
PART II: Neural Pathways for Color Vision
Three cone types are the basis of color vision (trichromacy):
- L cones - peak sensitivity at long wavelengths (~564 nm, red-sensitive)
- M cones - peak sensitivity at medium wavelengths (~534 nm, green-sensitive)
- S cones - peak sensitivity at short wavelengths (~420 nm, blue-sensitive)
Color information is coded as opponent-process signals along three parallel pathways from retina to V1:
- Red-green channel - signals the difference between L-cone and M-cone responses (L - M); carried by P ganglion cells → parvocellular LGN → deep layer 4C and blobs of V1
- Blue-yellow channel - signals S cones vs. the sum of L + M cones [S - (L+M)]; carried by bistratified (small) ganglion cells → koniocellular LGN → blobs of V1
- Luminance channel - signals the sum of L + M cone responses; carried by M ganglion cells → magnocellular LGN
From V1, color information projects from the blobs and layer 4C → V4/V8 in the ventral stream where the perception of color is generated. The exact mechanism by which V8 converts color input into conscious color sensation is not yet fully understood.
Color blindness is most commonly caused by absence or mutation of one cone type, typically:
- Absence of L cones (protanopia) or altered L pigment (protanomaly)
- Absence of M cones (deuteranopia) or altered M pigment (deuteranomaly)
- These are inherited as X-linked recessive traits, explaining the much higher prevalence in males (~8%) vs. females (~0.5%)
PART II: The Four Types of Eye Movements
The six extraocular muscles (innervated by CN III, IV, VI) share the same final common path for all eye movement types.
| Movement Type | Description | Function | Neural Control |
|---|
| Saccades | Sudden, rapid, jerky conjugate eye movements | Rapidly shift gaze from one object to another; bring new objects of interest onto the fovea; prevent adaptation of visual pathway that would occur with prolonged fixation | Programmed in the frontal eye fields (frontal cortex) and superior colliculi |
| Smooth pursuit movements | Slow, smooth tracking movements that follow a moving target | Keep a moving object's image stable on the fovea; prevent image blur during target motion | Depend on intact visual cortex; coordinated by the cerebellum |
| Vestibular movements | Compensatory eye movements driven by signals from the semicircular canals (vestibulo-ocular reflex) | Maintain stable visual fixation and keep images on the fovea as the head moves; prevent blurring during head rotation | Vestibular nuclei → CN III, IV, VI via the medial longitudinal fasciculus (MLF) |
| Convergence movements | Inward rotation of both eyes toward each other (disconjugate) | Bring both visual axes to bear on a near object; part of the near triad (with accommodation and pupillary constriction) | Midbrain convergence center; part of the accommodation response |
Strabismus arises when these movement systems are imbalanced. In children, the brain may suppress the image from the deviated eye to avoid diplopia, leading to amblyopia (lazy eye) if not corrected early.
Costanzo Physiology 7e, pp. 91-99 | Ganong's Review of Medical Physiology 26e, pp. 197-215