I now have all the material needed for a comprehensive answer. Let me compose the full response.
Neural Connections in the Cerebellar Cortex & Cerebellar Dysfunction
Part 1: Layers of the Cerebellar Cortex
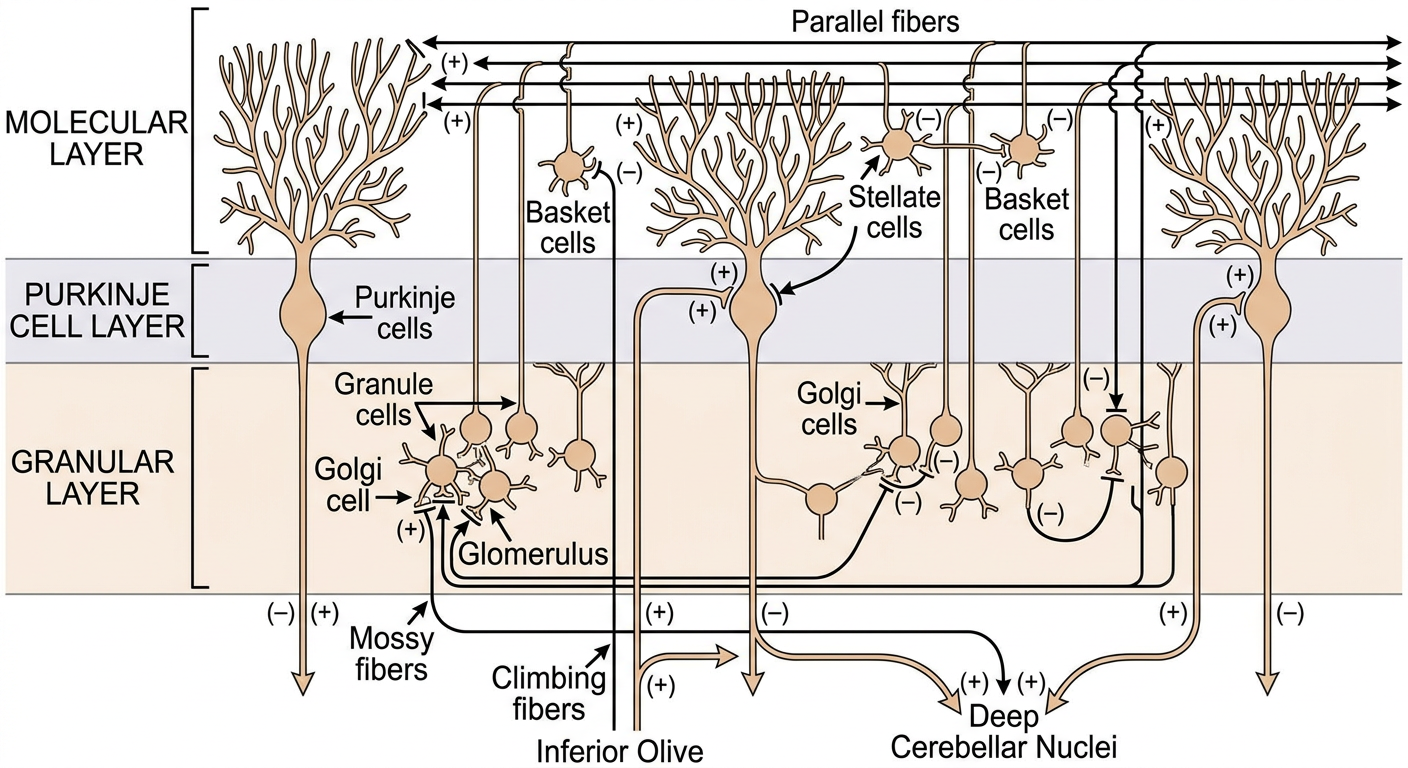
The cerebellar cortex has a remarkably uniform, three-layered structure throughout, containing five cell types. It is the only area of the CNS where the outermost layer is inhibitory (Purkinje cell output is entirely GABAergic).
Generated Diagram
Textbook Diagrams
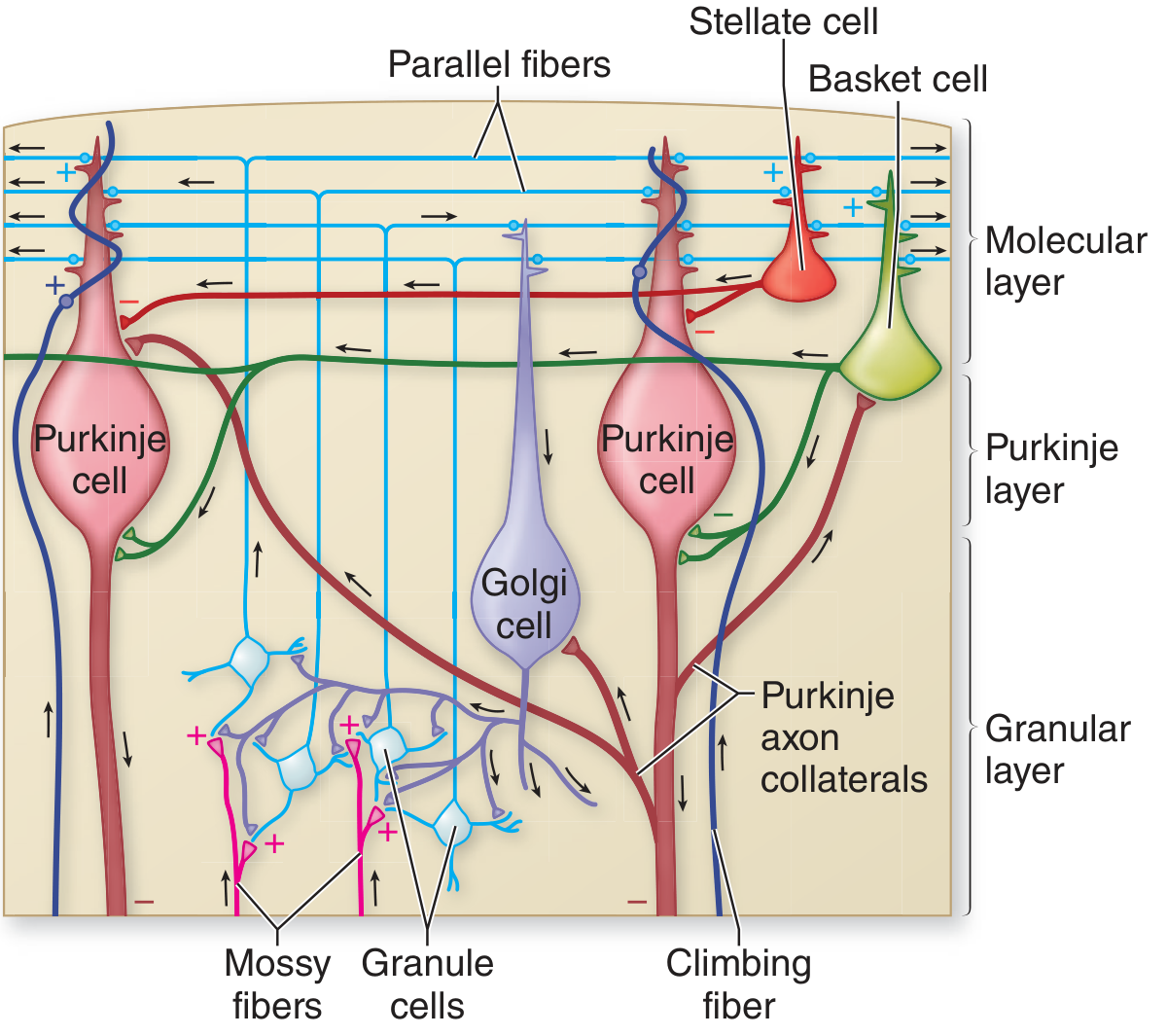
Fig. 3.36 - Costanzo Physiology (Structures of cerebellar cortex in cross-section):
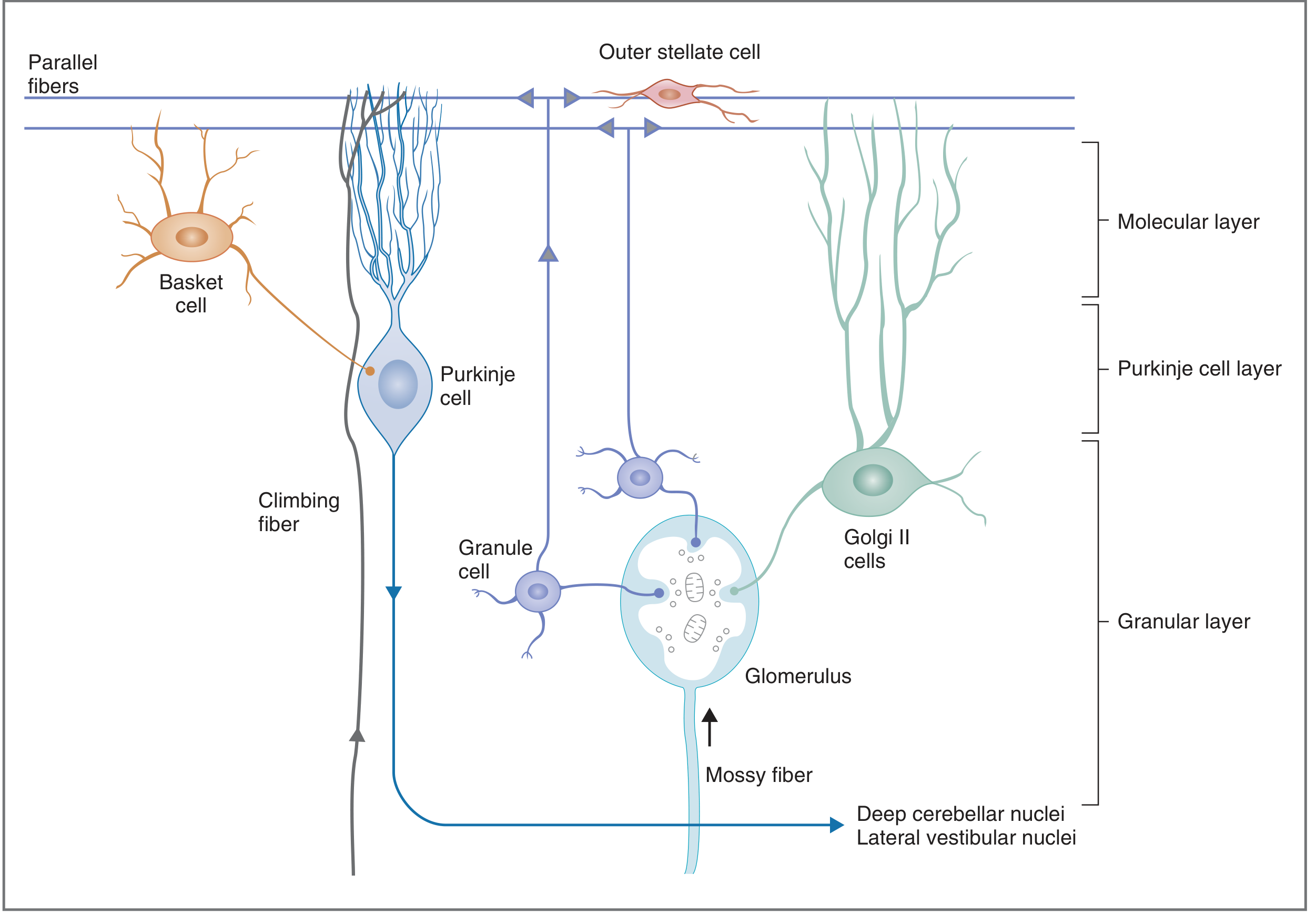
Fig. 5-6 - Adams & Victor (Physiologic organization showing excitatory/inhibitory connections):
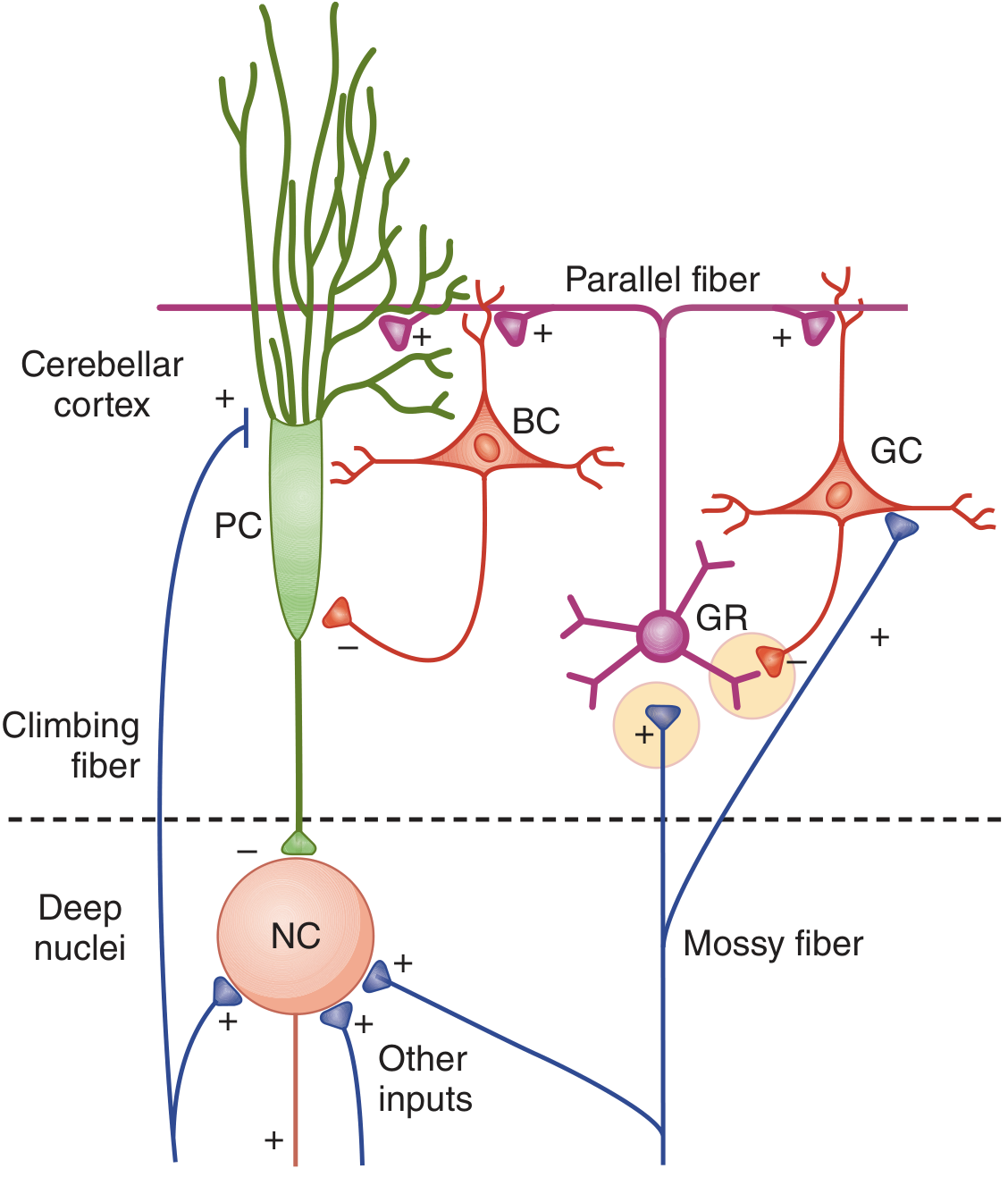
Fig. 12-18 - Ganong (Neural connections with BC, GC, GR, NC, PC labeling):
Part 2: Layers and Cell Types
1. Molecular Layer (outermost)
Contains:
- Outer stellate cells - inhibitory interneurons
- Basket cells - inhibitory interneurons
- Dendrites of Purkinje cells and Golgi II cells
- Parallel fibers - axons of granule cells running longitudinally
2. Purkinje Cell Layer (middle)
Contains:
- Purkinje cells - the sole OUTPUT neurons of the cerebellar cortex. Their output is ALWAYS inhibitory (GABA). Each Purkinje cell receives input from up to 1 million granule cells via parallel fibers, and from exactly one climbing fiber.
3. Granular Layer (innermost)
Contains:
- Granule cells - the only EXCITATORY neurons intrinsic to the cerebellar cortex (neurotransmitter: glutamate). Most numerous neurons in the entire CNS.
- Golgi type II cells - inhibitory interneurons
- Glomeruli - specialized synaptic complexes (see below)
Part 3: Inputs to the Cerebellar Cortex
Two major excitatory input systems reach the cerebellum. Both also send collateral branches directly to the deep cerebellar nuclei (bypassing the cortex).
A. Mossy Fiber System (majority of afferents)
| Feature | Detail |
|---|
| Origin | Vestibulocerebellar, spinocerebellar, and pontocerebellar tracts |
| Terminate on | Granule cell dendrites in glomeruli |
| Glomerulus | A specialized synaptic complex where mossy fiber terminals (+) and Golgi cell axon terminals (-) converge on granule cell dendrites. Enclosed in a glial sheath. |
| Effect on Purkinje cell | Weak but distributed - "simple spikes" (low amplitude, regular firing) via parallel fibers |
| Divergence | Each mossy fiber contacts many granule cells; each granule cell axon (parallel fiber) contacts up to 500 Purkinje cells |
Pathway: Mossy fiber → (+) Granule cell → Axon ascends → Bifurcates as Parallel fiber → (+) Purkinje cell dendrites → Purkinje cell axon → (-) Deep cerebellar nuclei
B. Climbing Fiber System
| Feature | Detail |
|---|
| Origin | Inferior olivary nucleus (medulla oblongata) |
| Terminate on | Directly on Purkinje cell dendrites, wrapping them |
| Convergence | Each Purkinje cell receives input from only ONE climbing fiber |
| Divergence | Each climbing fiber synapses on only 1-10 Purkinje cells |
| Effect | Powerful - produces complex spikes (large, high-frequency bursts) |
| Function | "Teaches" or conditions the Purkinje cell; plays a key role in cerebellar motor learning (error signal) |
Part 4: Intrinsic (Inhibitory) Interneurons and Their Connections
Basket Cells (Molecular Layer)
- Excited by parallel fibers (+)
- Send axons perpendicular to parallel fibers
- Form "basket"-like inhibitory (-) synapses on Purkinje cell bodies (and initial axon segment)
- Produce lateral (feed-forward) inhibition of adjacent Purkinje cells
- Function: Narrow the spatial extent of excitation (spatial signal sharpening)
Stellate Cells (Molecular Layer)
- Excited by parallel fibers (+)
- Inhibit (-) Purkinje cell dendrites
- Also produce feed-forward lateral inhibition
- Similar function to basket cells but act on dendrites, not soma
Golgi Cells (Granular Layer)
- Excited by: (a) mossy fiber collaterals, and (b) parallel fibers in the molecular layer
- Inhibit (-) granule cell dendrites at the glomerulus (feedback inhibition)
- Function: Shorten the duration of excitatory drive to granule cells - enhances temporal signal resolution
Part 5: Output of the Cerebellar Cortex
| Component | Description |
|---|
| Output neurons | Purkinje cells (only output cell) |
| Neurotransmitter | GABA (inhibitory) |
| Targets | Deep cerebellar nuclei (dentate, emboliform, globose, fastigial) and lateral vestibular nucleus |
| Net effect | Purkinje cell activity inhibits deep cerebellar nuclei |
| Deep cerebellar nuclei output | Always excitatory (to brainstem and thalamus → motor cortex) |
The deep cerebellar nuclei receive two opposing inputs:
- Excitatory (+) from mossy and climbing fiber collaterals (tonic excitation)
- Inhibitory (-) from Purkinje cell axons (phasic modulation)
This arrangement means the cerebellum modulates movement by varying the degree of Purkinje cell inhibition on the deep nuclei. When Purkinje cells are active, they reduce deep nuclei firing; when silenced (e.g., by climbing fiber complex spikes), they disinhibit the deep nuclei.
Summary Circuit (Text Diagram)
INPUTS:
Mossy fibers ──(+)──> Granule cells ──(+)──> Parallel fibers ──(+)──> Purkinje cell
↑ │ │
Golgi cell ──(-)──┘ └──(+)──> Basket cell ──(-)──> PC
↑ └──(+)──> Stellate cell──(-)──> PC
MF collateral (+) └──(+)──> Golgi cell ──(-)──> GrC
Climbing fibers (inf. olive) ──(+)──> Purkinje cell (complex spikes)
OUTPUTS:
Purkinje cell ──(-)──> Deep cerebellar nuclei ──(+)──> Thalamus → Motor cortex
↑
MF/CF collaterals (+) (tonic excitation)
Part 6: Physiological Basis of Clinical Features in Cerebellar Dysfunction
The cerebellum functions as a comparator: it receives a copy of the motor command (efference copy) and sensory feedback, and continuously corrects ongoing movements for accuracy of rate, range, force, and direction. Cerebellar dysfunction breaks this automatic coordination, forcing conscious cortical control of movements.
"Movements of my left arm are done subconsciously, but I have to think out each movement of the right arm." - Patient with right cerebellar hemisphere lesion (Gordon Holmes, as quoted in Kandel's Principles of Neural Science, 6th ed.)
1. Hypotonia
Clinical feature: Decreased muscle tone, "floppy" limbs, especially ipsilateral to the cerebellar lesion (most pronounced in proximal upper limb).
Physiological basis:
- The cerebellum maintains baseline excitatory drive to the motor cortex and spinal motor neurons via the dentate nucleus → thalamus → cortex pathway.
- Cerebellar lesions (especially dentate nucleus) reduce this tonic excitation, decreasing gamma motor neuron (fusimotor) drive to muscle spindles.
- Reduced spindle sensitivity → decreased stretch reflex activity → hypotonia.
- Pendular knee jerk: Normally, the muscle spindle reflex is damped by cerebellar-modulated opposing muscle contractions. With hypotonia, the leg swings freely like a pendulum after the patellar tap instead of returning to rest in 1-2 swings.
(Localization in Clinical Neurology, 8e - "hypotonia is probably the result of decreased fusimotor activity secondary to cerebellar injury, especially to the dentate nucleus")
2. Ataxia (Incoordination)
Clinical feature: Irregular, jerky, variable movements; errors in rate, range, direction, and force; wide-based gait; trunk sway.
Physiological basis:
- The cerebellum normally acts in feed-forward mode - predicting and pre-correcting movements before sensory feedback can arrive (sensory feedback is too slow for rapid movements).
- Cerebellar damage removes this predictive (feed-forward) control.
- Sequential agonist/antagonist contractions become mistimed: movements overshoot/undershoot (dysmetria), are broken into components (decomposition of movement), and force/velocity are poorly regulated.
- Gait ataxia results from failure to properly time weight transfer from one foot to the other and poor trunk stabilization; wide base is a compensatory strategy.
3. Dysmetria
Clinical feature: Inability to accurately gauge the distance/extent of a movement - finger-nose test shows past-pointing (hypermetria) or undershoot (hypometria).
Physiological basis:
- The cerebellum uses an internal model to predict how movements will unfold and programs the braking (deceleration) command in advance.
- Without cerebellar correction, the braking impulse to stop a movement is mistimed and insufficient.
- The limb overshoots (hypermetria) because the opposing muscle contraction arrives too late; or undershoots (hypometria) with excessive braking. Both result from lack of predictive motor programming.
- (Costanzo: "A limb may overshoot its target or stop before reaching its target")
4. Intention (Kinetic) Tremor
Clinical feature: Tremor that is absent at rest, appears and worsens as the limb approaches its target (end-point/terminal tremor). Distinguished from resting tremor of Parkinson's disease.
Physiological basis:
- Normally, the cerebellum dampens the oscillatory tendency of the limb by providing well-timed corrective signals.
- In cerebellar disease, the loss of predictive damping leads to a series of uncorrected oscillations - the limb overshoots, then the correction overshoots, and so on.
- The tremor worsens near the target because fine, precise corrections are most needed at endpoint and the cerebellar error-correction system fails most conspicuously here.
- The tremor is perpendicular to the direction of intended movement.
5. Dysdiadochokinesia
Clinical feature: Inability to perform rapid alternating movements (e.g., pronation-supination of forearm). Movements become slow, irregular, and arrhythmic.
Physiological basis:
- Rapid alternating movements require precise, rapid switching between agonist and antagonist muscle contractions.
- The cerebellum times the cessation of one movement and initiation of the next.
- With cerebellar damage, the timing signals that halt one movement and initiate its reverse are disrupted - each reversal requires a new, consciously planned motor program rather than being automatic.
- (Costanzo: "Ataxia can be exhibited as dysdiadochokinesis, in which a person is unable to perform rapid, alternating movements")
6. Rebound Phenomenon (Holmes' Rebound Test)
Clinical feature: When a patient is asked to flex the forearm against resistance that is suddenly released, the arm flies upward uncontrollably (failure to check).
Physiological basis:
- Normally, when resistance is released, the cerebellum anticipates the sudden unloading and preemptively activates the antagonist (triceps) before the agonist (biceps) can overshoot.
- In cerebellar disease, this predictive antagonist contraction is absent; the limb keeps moving after the resistance is removed.
- Reflects loss of anticipatory postural adjustment via cerebellar feed-forward control.
7. Gait Ataxia (Cerebellar Gait)
Clinical feature: Wide-based, staggering, irregular gait with variable step length and timing; inability to perform tandem walking (heel-to-toe).
Physiological basis:
- Effective gait requires precise timing of: (a) weight transfer, (b) axial postural corrections, (c) limb trajectory corrections.
- The cerebellum coordinates all these simultaneously in an automatic, predictive manner.
- Without cerebellar input, each step requires conscious, effortful control; the "automatic" walking program breaks down. Wide base is compensatory. Midline (vermis) lesions produce truncal ataxia and gait > limb ataxia.
8. Nystagmus
Clinical feature: Rhythmic, involuntary oscillation of the eyes with a slow phase and a fast corrective phase (especially gaze-evoked nystagmus with lateral gaze).
Physiological basis:
- The cerebellar flocculus/nodulus stabilizes gaze by maintaining appropriate vestibulo-ocular reflex (VOR) gain and controlling the velocity of smooth pursuit.
- The cerebellum acts as a "neural integrator" to hold the eyes in eccentric positions of gaze.
- Cerebellar damage impairs this integrator; the eyes drift back to midline (slow phase) and a corrective saccade brings them back to target (fast phase) - this repetition produces nystagmus.
- Ocular dysmetria (overshoot saccades) occurs by the same mechanism as limb dysmetria.
9. Scanning (Ataxic) Dysarthria
Clinical feature: Slurred, irregular, slow speech with variable rhythm, volume and pitch; sometimes explosive, sometimes poorly articulated.
Physiological basis:
- Coordination of the 100+ muscles required for speech (respiration, larynx, palate, tongue, lips) requires the same precise timing and feed-forward control as limb movements.
- Cerebellar damage disrupts the timing of these muscular contractions; articulatory movements are irregular, poorly timed, and inaccurate - producing "scanning speech" where each syllable is produced with equal, unnatural emphasis.
- In acute/severe damage, speech may also be explosive (sudden bursts).
10. Decomposition of Movement (Dyssynergia)
Clinical feature: Multi-joint movements are broken into their component single-joint movements, performed sequentially rather than smoothly together.
Physiological basis:
- The cerebellum coordinates synergistic muscle groups - muscles that must act together to produce smooth multi-joint motion.
- At joints not directly controlled, reactive forces from adjacent joint movement are normally anticipated (predicted) and preemptively compensated by the cerebellum.
- Cerebellar damage removes this inter-joint prediction; the patient compensates by simplifying movement into single-joint components (though this is only partially effective). (Kandel: "patients often have difficulty holding the shoulder steady while moving the elbow, a deficit thought to be due to poor predictions of how the movement at the elbow mechanically affects the shoulder")
Summary Table: Clinical Features and Physiological Basis
| Clinical Feature | Physiological Basis |
|---|
| Hypotonia | ↓ Dentate nucleus output → ↓ fusimotor (gamma) drive → ↓ spindle sensitivity; pendular reflexes |
| Ataxia | Loss of feed-forward predictive motor control; errors in rate/range/force/direction |
| Dysmetria | Failure of braking impulse timing; absent predictive antagonist activation |
| Intention tremor | Loss of oscillation-damping; uncorrected sequential correction overshoots |
| Dysdiadochokinesia | Impaired timing of agonist-antagonist switching |
| Rebound phenomenon | Absent anticipatory antagonist contraction when resistance released |
| Gait ataxia | Failure of automatic weight transfer timing; loss of predictive postural adjustment |
| Nystagmus | Failure of gaze-holding neural integrator (flocculus); impaired VOR gain |
| Scanning dysarthria | Incoordination of speech musculature timing |
| Decomposition | Loss of inter-joint force prediction; synergistic muscle grouping disrupted |
Key Clinical Principle: Ipsilateral Localization
Unlike cerebral cortex lesions (which cause contralateral deficits), cerebellar hemisphere lesions cause ipsilateral signs because:
- The cerebellum exerts ipsilateral control via the cerebellocortical pathway which crosses twice (once in the superior cerebellar peduncle and once in the corticopontine tract), returning control to the ipsilateral side.
- Midline (vermis) lesions predominantly cause truncal ataxia and gait disturbance.
- Lateral hemisphere lesions cause limb incoordination.
- Flocculonodular lesions cause vestibular signs (nystagmus, vertigo).
Sources:
- Costanzo Physiology, 7th ed., pp. 113-115
- Adams & Victor's Principles of Neurology, 12th ed., pp. 125-126
- Kandel's Principles of Neural Science, 6th ed., pp. 956-957
- Ganong's Review of Medical Physiology, 26th ed., pp. 255-256
- Localization in Clinical Neurology, 8th ed., pp. 967-968
- Neuroanatomy through Clinical Cases, 3rd ed., pp. 731-732