Describe and differentiate the properties of skeletal, cardiac & smooth muscle.
Reading File
Finding Sources
Finding Sources
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
I now have all the content needed for a thorough, well-structured answer. Here is the complete comparison:
Skeletal, Cardiac & Smooth Muscle: Properties and Differences
The three types of muscle in vertebrates all convert ATP chemical energy into mechanical work, but they differ profoundly in structure, control, location, and contractile mechanism. Skeletal and cardiac muscle are striated due to the ordered parallel alignment of their contractile filaments into sarcomeres. Smooth muscle lacks striations because its contractile fibrils are arranged more randomly - which also means it can generate force in all directions (like an inflating balloon), while striated muscles generate force in one dimension only (like a coil spring).
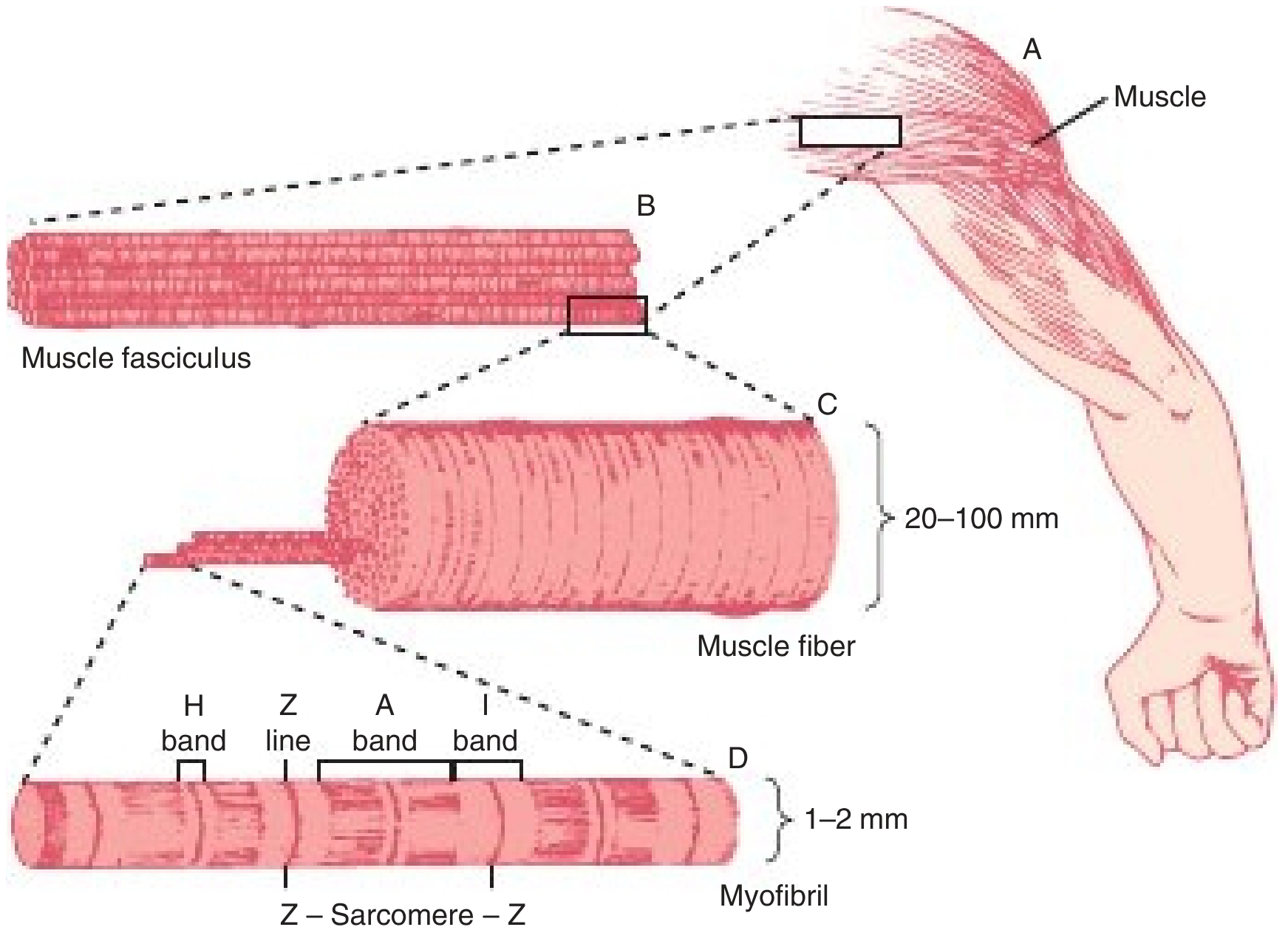
Structure of voluntary muscle - Harper's Illustrated Biochemistry, 32nd Ed, p. 630
1. Structural Features
| Feature | Skeletal | Cardiac | Smooth |
|---|---|---|---|
| Cell size | Large, 10-100 µm diameter; up to 100 cm long | Short/narrow, 10-100 µm diameter, 80-100 µm long | Fusiform, 0.2-2 µm diameter, 20-200 µm long |
| Nucleus | Many, peripheral | Single, central (with perinuclear organelle zone) | Single, central |
| Striations | Present | Present | Absent |
| Sarcomere | Present (1,500-2,300 nm, Z-line to Z-line) | Present | Absent |
| T tubules | At A-I junction; triad (one T tubule + two terminal cisternae); 2 per sarcomere | At Z lines; diad (one T tubule + small terminal cisternae); 1 per sarcomere | Absent - replaced by caveolae and cytoplasmic vesicles |
| Cell-to-cell junctions | None (each fiber is independent) | Intercalated discs containing fasciae adherentes, desmosomes, and gap junctions | Gap junctions (nexus) |
| Special structures | Well-developed sarcoplasmic reticulum (SR) and T tubules | Intercalated discs, Purkinje fibers | Dense bodies (α-actinin rich); desmin/vimentin intermediate filaments; caveolae |
| Connective tissue | Epimysium, perimysium, endomysium | Endomysium, subendocardial/subpericardial CT | Endomysium, sheaths, bundles |
- Histology: A Text and Atlas, p. 872
- Guyton & Hall Textbook of Medical Physiology
Key structural point - intercalated discs: Cardiac muscle cells are connected end-to-end by intercalated discs, which contain gap junctions allowing rapid ion diffusion. This makes the entire cardiac muscle a functional syncytium - an action potential spreads from cell to cell so the atria contract as one unit and the ventricles as another. Skeletal muscle has no such electrical coupling between fibers.
2. Location
| Type | Location |
|---|---|
| Skeletal | Muscles of the skeleton; visceral striated muscle (tongue, esophagus, diaphragm) |
| Cardiac | Heart wall; portions of superior/inferior vena cavae and pulmonary veins |
| Smooth | Walls of blood vessels, hollow organs (GI tract, bladder, uterus), bronchi, iris |
3. Control (Voluntary vs. Involuntary)
- Skeletal - voluntary, under conscious control via the somatic nervous system
- Cardiac - involuntary; has intrinsic automaticity (pacemaker cells in the SA node), modulated (but not initiated) by the autonomic nervous system and circulating hormones
- Smooth - involuntary; controlled by the autonomic nervous system, local paracrine factors, hormones, mechanical stretch, and intrinsic pacemaker activity in some tissues
4. Mechanism of Excitation (Excitation-Contraction Coupling)
| Property | Skeletal | Cardiac | Smooth |
|---|---|---|---|
| Excitation mechanism | Neuromuscular junction; ACh activates nicotinic ACh receptor | Pacemaker depolarization spread electrotonically via gap junctions | Synaptic transmission, agonist-activated receptors, electrical coupling, pacemaker potentials |
| Electrical activity | Action potential spikes | Action potential plateaus (prolonged) | Spikes, plateaus, graded membrane potentials, slow waves |
| EC coupling | L-type Ca²⁺ channel (Cav1.1) in T-tubule mechanically gates RYR1 in SR (no Ca²⁺ entry needed) | Ca²⁺ entry via Cav1.2 triggers Ca²⁺-induced Ca²⁺ release (CICR) via RYR2 in SR | Ca²⁺ entry via Cav1.2; Ca²⁺- or IP₃-mediated release via RYR3 or IP₃R from SR; also store-operated Ca²⁺ entry (SOCs via Orai/TRP channels) |
| Ca²⁺ sensor | Troponin C | Troponin C | Calmodulin (CaM) |
| Terminator of contraction | ACh breakdown by acetylcholinesterase; SR Ca²⁺ uptake | Action potential repolarization; SR Ca²⁺ uptake | SR Ca²⁺ uptake; sarcolemmal Ca²⁺ extrusion |
- Medical Physiology (Boron & Boulpaep), Table 9-3, p. 395
Key EC coupling distinction: In skeletal muscle, the DHP receptor in the T-tubule directly gates the ryanodine receptor in the SR by mechanical (conformational) coupling - no actual Ca²⁺ influx is required. In cardiac muscle, Ca²⁺ must enter through L-type channels first to trigger CICR from the SR - this is why cardiac contractility is sensitive to extracellular Ca²⁺ levels. Smooth muscle uses calmodulin rather than troponin as its Ca²⁺ sensor.
5. Contractile Mechanism
Skeletal & Cardiac (sarcomere-based sliding filament):
- Thick filaments (myosin) and thin filaments (actin + troponin + tropomyosin) are arranged in sarcomeres
- Ca²⁺ binds troponin C → conformational change in tropomyosin → myosin binding sites on actin exposed → cross-bridge cycling
- Force is proportional to filament overlap
- Thick filaments are bipolar (antiparallel)
Smooth muscle:
- No sarcomeres, no troponin
- Thin filaments (actin + tropomyosin + caldesmon + calponin) anchor to dense bodies in the cytoplasm and dense bands at the sarcolemma
- Thick filaments are side-polar (not bipolar) - unique arrangement allowing greater shortening
- Contraction initiated by Ca²⁺-calmodulin activation of myosin light-chain kinase (MLCK), which phosphorylates myosin regulatory light chains, enabling cross-bridge cycling
- Smooth muscle is specialized for slow, sustained, energy-efficient contractions ("latch" state)
6. Regeneration and Repair
| Type | Repair capacity |
|---|---|
| Skeletal | Good - satellite cells (Pax7⁺ myogenic stem cells left from fetal development) are activated by injury, co-express Pax7 + MyoD, and regenerate muscle fibers |
| Cardiac | Poor - injury (e.g. MI) causes myocyte death replaced by fibrous connective tissue (scar); minimal endogenous regeneration |
| Smooth | Good - smooth muscle cells retain the ability to divide and maintain/increase in number; major remodeling occurs physiologically (e.g. uterine hypertrophy during pregnancy) |
- Histology: A Text and Atlas, p. 878
7. Functional Characteristics at a Glance
| Property | Skeletal | Cardiac | Smooth |
|---|---|---|---|
| Speed of contraction | Fast to slow (fiber-type dependent) | Intermediate | Slow |
| Fatigue | Fatigable (especially fast fibers) | Highly fatigue-resistant | Fatigue-resistant |
| Automaticity | None | Yes (pacemaker cells) | Some tissues (e.g. GI, ureter) |
| Tetanus possible? | Yes | No (long refractory period prevents tetanus) | Some forms of sustained contraction |
| Length-tension relationship | Yes | Yes (Frank-Starling law) | Modified - wide range of lengths before tension falls |
| Fiber arrangement effect | Unidirectional force | Unidirectional (but helical layering aids ventricular torsion) | Multi-directional force (all planes) |
Summary
The three muscle types represent an evolutionary spectrum. Smooth muscle is the most primitive, found in all organs and vessels, controlled by autonomic/hormonal signals, and capable of sustained low-energy contraction. Cardiac muscle evolved in the direction of skeletal muscle (acquiring sarcomeres and striations) while retaining characteristics of smooth muscle (central nuclei, gap junctions, automaticity). Skeletal muscle is the most specialized for rapid, powerful, precisely controlled voluntary movement. The key mechanistic distinction is in their Ca²⁺ sensing: skeletal and cardiac muscle use troponin C, while smooth muscle uses calmodulin - a difference that underpins their different pharmacological sensitivities and regulation.
Sources:
- Histology: A Text and Atlas with Correlated Cell and Molecular Biology, pp. 872-878
- Medical Physiology (Boron & Boulpaep), Table 9-3, p. 395
- Harper's Illustrated Biochemistry, 32nd Ed, pp. 630-631
- Guyton & Hall Textbook of Medical Physiology
This is a shared conversation. Sign in to Orris to start your own chat.