Physiology of Hearing: Outer, Middle, and Inner Ear. Physiology of Vision: Visual Pathways: Functional anatomy of the eye. Central visual pathways from retina to visual cortex. Explain each and every point in detail
Reading File
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Reading File
Finding Sources
Finding Sources
Finding Sources
Loading Image
Loading Image
Loading Image
Loading Image
I have more than enough detailed textbook content from Ganong's to write a comprehensive answer. Let me now compile the complete, detailed response.
Physiology of Hearing: Outer, Middle, and Inner Ear
PART 1: PHYSIOLOGY OF HEARING
Sound Waves - The Physical Basis of Hearing
Sound is the sensation produced when longitudinal vibrations of molecules in the external environment strike the tympanic membrane. These movements are expressed as changes in pressure over time, called sound waves.
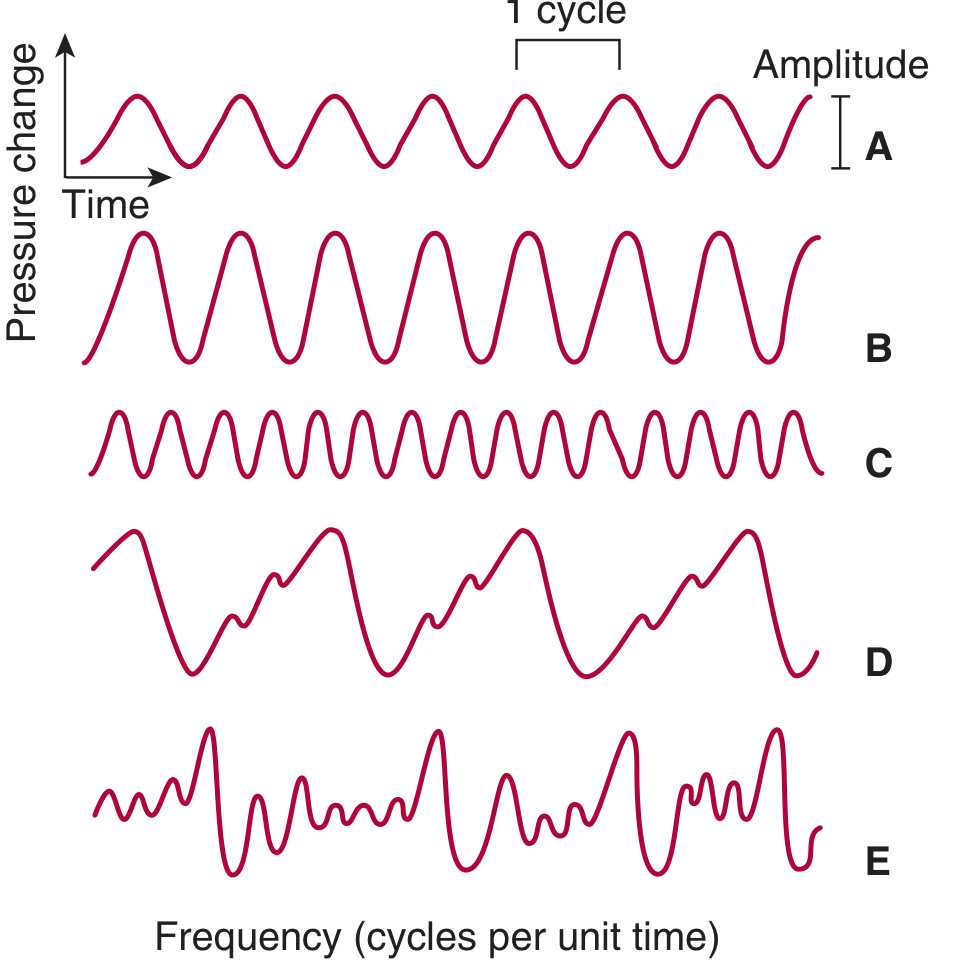
A = pure tone; B = greater amplitude (louder); C = same amplitude but higher frequency (higher pitch); D = complex but regularly repeated waveform (musical); E = aperiodic, irregular waves (noise)
Key physical properties:
- Sound travels through air at 344 m/s at 20°C at sea level (faster with temperature/altitude increase; 1450 m/s in fresh water)
- Loudness correlates with amplitude; Pitch correlates with frequency (Hz)
- Musical sounds = repeating waveform patterns; Noise = aperiodic, non-repeating vibrations
- Timbre (quality) allows us to distinguish instruments at the same pitch - due to harmonic overtones superimposed on the primary frequency
The Decibel Scale:
- Amplitude is measured logarithmically in decibels (dB)
- 0 dB = standard reference pressure of 0.000204 dyne/cm² (human auditory threshold)
- The 0-140 dB range represents a 10-million-fold variation in sound pressure
- 30 dB = whisper; 60-80 dB = conversation; 90-110 dB = subway/chain saw; 120-160 dB = painful/damaging
- Prolonged exposure >85 dB can cause hearing loss
Human Audibility:
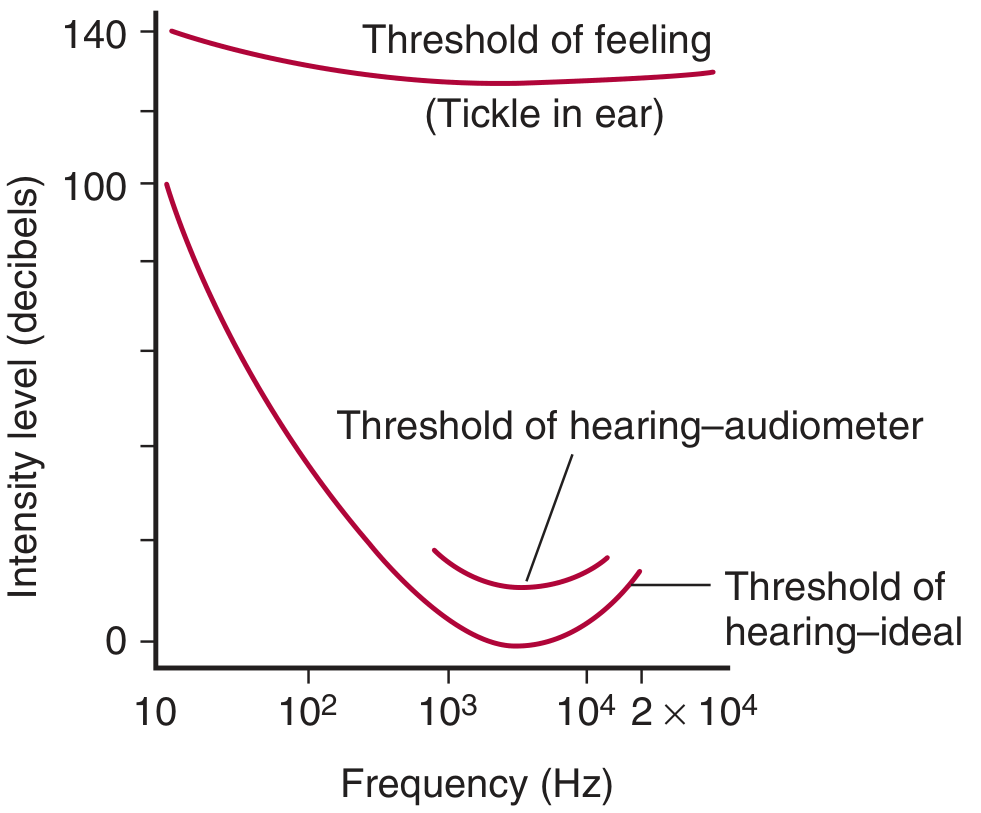
- Audible range: 20 to 20,000 Hz
- Greatest sensitivity: 1000-4000 Hz range
- Average male voice: ~120 Hz; Average female voice: ~250 Hz
- ~2000 distinguishable pitches in untrained individuals; trained musicians discriminate far more
THE OUTER EAR
Anatomy and Function
The outer ear consists of:
- Pinna (Auricle) - the external cartilaginous structure
- External auditory meatus (EAM) - the canal leading to the tympanic membrane
Functions of the Outer Ear:
-
Sound collection and funneling - The pinna collects sound waves from the environment and directs them down the external auditory canal toward the tympanic membrane.
-
Sound localization - The shape and ridges of the pinna introduce direction-dependent transformations (especially for vertical localization). The asymmetric shape causes reflections that differ depending on whether sound comes from above, below, front, or behind - this is called the Head-Related Transfer Function (HRTF).
-
Resonance amplification - The external auditory canal (~2.5 cm long) acts as a resonator. It resonates at approximately 3000-4000 Hz, amplifying sounds in this range by about 10-15 dB. This is why humans are most sensitive in this frequency range.
-
Protection - The canal's orientation, cerumen (earwax), and the tympanic membrane at its end protect the middle ear from physical and infectious agents.
The Tympanic Membrane (Eardrum):
- A thin, semi-transparent, cone-shaped membrane separating the outer and middle ear
- The apex of the cone (umbo) is attached to the handle of the malleus
- When sound waves strike it, it vibrates in unison with the sound wave frequency
- Area: ~85 mm²; the effective vibrating area is about 55 mm²
THE MIDDLE EAR
Anatomy
The middle ear is an air-filled cavity (tympanic cavity) containing three ossicles:
- Malleus (hammer) - attached to the tympanic membrane at the umbo
- Incus (anvil) - connects malleus to stapes
- Stapes (stirrup) - footplate sits in the oval window of the cochlea
The middle ear communicates with the nasopharynx via the Eustachian tube, which equalizes pressure on both sides of the tympanic membrane.
The Impedance-Matching Function (Core Role of the Middle Ear)
The middle ear solves a fundamental physics problem: sound travels efficiently through air but loses ~99.9% of its energy (30 dB loss) when it tries to cross from an air medium to a fluid medium (like the perilymph in the cochlea). The middle ear acts as an impedance transformer to overcome this mismatch.
This is achieved by three mechanisms:
1. Area ratio (most important factor - ~25-30 dB gain):
- Tympanic membrane effective area: ~55 mm²
- Stapes footplate area: ~3.2 mm²
- Ratio = ~17:1
- Pressure = Force/Area; since force is transmitted from a large area to a small area, pressure is amplified ~17-fold (= ~25 dB gain)
2. Lever action of the ossicular chain (~2-3 dB gain):
- The malleus handle is longer than the long process of the incus
- Handle of malleus : long process of incus = approximately 1.3:1
- This lever action provides an additional mechanical advantage of about 1.3-fold
3. Buckling motion of the tympanic membrane:
- The curved shape of the tympanic membrane doubles the force at the center (umbo), providing a further amplification
Combined effect: ~25-30 dB gain, almost exactly compensating for the energy that would otherwise be lost at the air-fluid interface.
The Stapedius and Tensor Tympani Reflexes (Acoustic Reflex)
Two tiny muscles in the middle ear protect it from loud sounds:
- Tensor tympani (innervated by CN V3 - trigeminal nerve): pulls the malleus medially, tensing the tympanic membrane
- Stapedius (innervated by CN VII - facial nerve): pulls the stapes posteriorly, reducing its vibration amplitude
When sounds >70-80 dB are detected, both muscles contract reflexively - the acoustic (stapedial) reflex. This:
- Stiffens the ossicular chain
- Reduces transmission of low-frequency sounds by ~10-15 dB
- Protects the cochlea from damage by sustained loud sounds
- The reflex latency is 25-150 ms, so it does NOT protect against sudden loud impulses (e.g., gunshots)
THE INNER EAR (COCHLEA)
Gross Structure of the Cochlea
The cochlea is a snail-shaped, fluid-filled bony labyrinth coiled 2.5 turns around a central bony pillar called the modiolus. It is divided longitudinally into three fluid-filled compartments by two membranes:
- Scala vestibuli (upper compartment) - filled with perilymph; communicates with the middle ear at the oval window
- Scala media (cochlear duct) - middle compartment - filled with endolymph (high K⁺, low Na⁺); contains the organ of Corti
- Scala tympani (lower compartment) - filled with perilymph; communicates with the middle ear at the round window
The scala vestibuli and scala tympani communicate at the apex of the cochlea through a small opening called the helicotrema.
Separating membranes:
- Reissner's membrane (vestibular membrane) - separates scala vestibuli from scala media
- Basilar membrane - separates scala media from scala tympani; critically important for frequency analysis
Perilymph vs. Endolymph
| Property | Perilymph | Endolymph |
|---|---|---|
| Location | Scala vestibuli + tympani | Scala media |
| [Na⁺] | ~140 mEq/L (like ECF) | ~1 mEq/L |
| [K⁺] | ~5 mEq/L | ~150 mEq/L (like ICF) |
| Resting potential | 0 mV | +80 mV (endocochlear potential) |
| Produced by | Perilymphatic space | Stria vascularis |
The endocochlear potential (+80 mV) maintained by the stria vascularis creates a large electrochemical gradient that is essential for hair cell mechanotransduction.
The Basilar Membrane and Frequency Analysis (Place Theory / Tonotopy)
The basilar membrane is the key structure for analyzing the frequency content of sound. It is not uniform along its length:
| Location | Width | Stiffness | Best Frequency |
|---|---|---|---|
| Base (near oval window) | Narrow | Stiff | High frequencies (15,000-20,000 Hz) |
| Apex (near helicotrema) | Wide | Flaccid | Low frequencies (20-200 Hz) |
When the stapes vibrates at the oval window, it sets up a traveling wave that propagates along the basilar membrane from base to apex. Each frequency causes maximum displacement at a specific point - this is the basis of Bekesy's Place Theory (confirmed by Georg von Bekesy, Nobel Prize 1961):
- The point of maximum displacement encodes the frequency of the incoming sound
- High-frequency sounds cause peak displacement at the base
- Low-frequency sounds travel further, peaking at the apex
This tonotopic organization is preserved all the way to the auditory cortex.
The Organ of Corti
The organ of Corti sits on the basilar membrane within the scala media and is the actual mechanosensory transducer.
Structure:
- Runs the entire length of the cochlea (~35 mm)
- Contains about 15,000-16,000 hair cells arranged in rows
- Covered by the tectorial membrane (a gelatinous shelf projecting from the modiolus)
Hair cells - two types:
Inner Hair Cells (IHCs):
- Single row, ~3,500 cells total
- Responsible for ~95% of afferent signals to the auditory nerve
- The primary sensory transducers
- Shaped like a flask; tips have 40-70 stereocilia arranged in a V or U pattern
Outer Hair Cells (OHCs):
- Three rows, ~12,000 cells total
- Primarily electromotile (cochlear amplifiers) - they actively contract and elongate
- Driven by a motor protein called prestin in their lateral membrane
- Amplify the traveling wave by up to 40-50 dB, especially at low sound levels
- Receive mostly efferent innervation (olivocochlear bundle from superior olivary complex)
- Loss of OHCs = high-frequency sensorineural hearing loss
Mechanotransduction - How Hair Cells Convert Sound to Electrical Signals
This is the fundamental step of hearing:
-
Basilar membrane deflects upward (toward tectorial membrane) due to traveling wave
-
Stereocilia deflect - the tips of the IHC stereocilia are embedded in (or closely apposed to) the tectorial membrane. When the basilar membrane moves, a shearing motion occurs between the basilar membrane and the tectorial membrane, deflecting the stereocilia.
-
Tip links connect the tips of shorter stereocilia to the sides of taller ones. When stereocilia deflect toward the tallest row, tip links pull open mechanosensitive K⁺ channels (MET channels - MYO7A/TMIE complex at the tips).
-
K⁺ influx - Because endolymph is rich in K⁺ (+150 mEq/L) and the inside of the hair cell is negative (~-70 mV), K⁺ rushes INTO the hair cell through the MET channels when they open. This is unusual - most K⁺ channels cause K⁺ to leave cells.
-
Depolarization - The K⁺ influx depolarizes the hair cell membrane.
-
Ca²⁺ influx and vesicle fusion - Depolarization opens voltage-gated Ca²⁺ channels at the base of the IHC, triggering exocytosis of glutamate at the ribbon synapse onto the afferent auditory nerve fiber (spiral ganglion cell type I).
-
Action potential generation - The released glutamate activates AMPA receptors on the afferent auditory nerve, generating action potentials in the cochlear nerve (CN VIII).
-
Repolarization - K⁺ exits through basolateral K⁺ channels (KCNQ4) into perilymph of scala tympani, then is recycled back to endolymph via supporting cells and stria vascularis.
When stereocilia deflect AWAY from tallest row - MET channels close → hyperpolarization → reduced neurotransmitter release → reduced firing.
Auditory Nerve Coding
- Frequency coding: Place code (which hair cells are activated) + temporal coding (phase-locking of nerve firing to the stimulus frequency up to ~4000 Hz)
- Intensity coding: Increased sound intensity causes:
- Greater amplitude of basilar membrane displacement → more hair cells activated (broader tuning)
- Higher rate of firing in individual nerve fibers
- Recruitment of high-threshold nerve fibers
Each auditory nerve fiber has a characteristic frequency (CF) - the frequency at which it is most sensitive. The tuning is sharp (narrow bandwidth) due to active OHC amplification.
THE AUDITORY PATHWAY (Central Processing)
Once the cochlear nerve fibers fire, the signal travels through a multi-synaptic relay:
1. Cochlear Nerve (CN VIII, Auditory Portion)
- ~30,000 afferent fibers from spiral ganglion neurons (95% type I from IHCs; 5% type II from OHCs)
- Fibers travel in the internal auditory canal → enter brainstem at the pontomedullary junction
2. Cochlear Nuclei (Medulla)
- ALL cochlear nerve fibers synapse in the dorsal and ventral cochlear nuclei on the ipsilateral side
- This is the only obligatory synapse where tonotopy is faithfully preserved
- Second-order neurons show sharper frequency tuning than first-order neurons - due to lateral inhibition in the brainstem
3. Superior Olivary Complex (SOC) - First Binaural Processing
- Most fibers from cochlear nuclei cross the midline via the trapezoid body to the contralateral SOC
- Some fibers synapse ipsilaterally
- The SOC is the first site of binaural convergence - crucial for sound localization
- Medial Superior Olive (MSO): detects interaural TIME differences (ITDs) - important at low frequencies (<3000 Hz)
- Lateral Superior Olive (LSO): detects interaural LEVEL differences (ILDs) - important at high frequencies (>3000 Hz)
- Efferent olivocochlear fibers originate here and project back to OHCs
4. Inferior Colliculus (Midbrain Tectum)
- Most ascending fibers synapse here via the lateral lemniscus
- Critical integration center for spatial hearing and complex sound analysis
- Both crossed and uncrossed projections converge
- Strong projections control the acoustic startle reflex (via reticular formation)
5. Medial Geniculate Body (MGB) - Thalamus
- Thalamic relay for all ascending auditory information
- Strictly tonotopically organized
- Projects to the auditory cortex via the auditory radiation
6. Primary Auditory Cortex (A1) - Heschl's Gyri
- Located on the superior temporal gyrus (transverse temporal gyri, Heschl's convolutions) - Brodmann areas 41 and 42
- Strictly tonotopic: low frequencies anterolaterally, high frequencies posteromedially
- Most neurons respond to both ears (binaural)
- Strips of cells activated by contralateral input are inhibited by ipsilateral input
Hemispheric Specialization
- Left auditory cortex / Wernicke area (Brodmann 22): specialized for language/speech processing; far more active during language tasks
- Right auditory cortex: specialized for melody, pitch, sound intensity, and prosody
- The auditory cortex shows remarkable plasticity: in prelingually deaf individuals who learn sign language, sign language activates auditory association areas; individuals blind from early life show better sound localization than sighted people
PART 2: PHYSIOLOGY OF VISION
FUNCTIONAL ANATOMY OF THE EYE
Overview
The eye is an optical instrument that focuses images of the external world onto a layer of photoreceptors at the back of the eye (the retina), which then converts light energy into electrical signals.
The Three Concentric Coats (Tunics) of the Eyeball
1. Outer fibrous coat:
- Cornea (anterior 1/6): transparent, avascular, the primary refracting surface of the eye. It accounts for approximately +43 diopters of the total ~+60 diopter refracting power of the eye. Nourished by aqueous humor and limbal vessels. Has five layers: epithelium, Bowman layer, stroma, Descemet membrane, endothelium.
- Sclera (posterior 5/6): opaque, white, fibrous coat that gives the eye its shape and provides attachment for extraocular muscles.
2. Middle vascular coat (uvea):
- Choroid: highly vascular, pigmented layer between sclera and retina; supplies the outer retinal layers (photoreceptors) with oxygen and nutrients
- Ciliary body: produces aqueous humor and contains the ciliary muscle (smooth muscle) which controls accommodation via changes in lens shape
- Iris: pigmented diaphragm with a central aperture (the pupil); contains:
- Sphincter pupillae (circular smooth muscle, parasympathetic CN III): pupil constriction (miosis)
- Dilator pupillae (radial smooth muscle, sympathetic): pupil dilation (mydriasis)
3. Inner nervous coat:
- Retina: discussed in detail below
Key Optical Structures
The Lens:
- Biconvex, transparent, avascular, elastic structure
- Enclosed in the lens capsule and suspended by zonule fibers (suspensory ligaments of Zinn) connecting to the ciliary body
- Contributes approximately +20 diopters of refracting power (variable, +0 to +16 D more via accommodation)
- Composed of lens fibers (elongated epithelial cells filled with crystallins)
- With age, the lens loses elasticity → presbyopia
Accommodation:
For near vision, the ciliary muscle contracts → zonule fibers loosen → lens becomes more convex (higher power) → closer objects brought into focus. For far vision: ciliary muscle relaxes → zonules tighten → lens flattens.
The Cornea-Lens System:
- Far point (emmetropia): parallel rays focus exactly on the retina without accommodation
- Myopia (short-sight): eyeball too long OR cornea too curved → focus in front of retina
- Hyperopia (long-sight): eyeball too short → focus behind retina
- Astigmatism: non-spherical cornea/lens → different focal points in different meridians
Aqueous Humor:
- Produced by ciliary processes via active secretion and ultrafiltration
- Flows from posterior chamber → through pupil → anterior chamber → drains via trabecular meshwork into Schlemm's canal → venous circulation
- Maintains intraocular pressure (normal: 10-21 mmHg)
- Obstruction of drainage → glaucoma
Vitreous Humor:
- Gel-like substance filling the posterior segment
- Composed of water, collagen fibrils, and hyaluronic acid
- Not replaced; provides structural support and maintains the retina in apposition to the RPE
The Retina - Detailed Structure
The retina is a ~0.5 mm thick sheet of neural tissue. It has 10 layers (from inside/vitreal to outside/scleral):
- Inner limiting membrane (Muller cell end-feet)
- Nerve fiber layer (ganglion cell axons)
- Ganglion cell layer
- Inner plexiform layer (synapse between bipolar/amacrine and ganglion cells)
- Inner nuclear layer (bipolar, amacrine, horizontal cells)
- Outer plexiform layer (synapse between photoreceptors and bipolar/horizontal cells)
- Outer nuclear layer (photoreceptor nuclei)
- Outer limiting membrane
- Photoreceptor layer (rods and cones)
- Retinal Pigment Epithelium (RPE)
Key Retinal Regions:
Fovea centralis:
- ~0.35 mm diameter pit at the center of the macula
- Contains ONLY cones (~35,000 in the central fovea/foveola)
- The overlying layers are displaced laterally (foveal pit) so light reaches cones directly with minimal scattering
- Provides highest visual acuity and color vision
- Each foveal cone connects to ONE ganglion cell (1:1 ratio) → maximum spatial resolution
Macula lutea:
- ~5 mm region surrounding the fovea
- Contains yellow pigment (xanthophyll: lutein + zeaxanthin) that absorbs blue/UV light and protects against phototoxic damage
Optic disc (blind spot):
- Where the optic nerve exits the eye; ~1.5 mm diameter
- Contains NO photoreceptors → creates a physiological blind spot (~15° temporal from the fixation point)
- Also where the central retinal artery and vein enter/exit
Peripheral retina:
- Dominated by rods (low light, motion detection, no fine detail)
- Multiple rods converge onto single ganglion cells (many-to-one convergence) → high sensitivity but low acuity
Photoreceptors - Rods and Cones
| Feature | Rods | Cones |
|---|---|---|
| Number | ~120 million | ~6-7 million |
| Distribution | Peripheral retina | Concentrated in fovea |
| Light sensitivity | Scotopic (dim light) | Photopic (bright light) |
| Wavelength peaks | ~498 nm (rhodopsin) | S: 420 nm (blue); M: 531 nm (green); L: 558 nm (red) |
| Acuity | Low | High |
| Color | No | Yes (3 types: S, M, L cones) |
| Dark current | Large | Smaller |
Structure of Photoreceptors:
Each photoreceptor has:
- Outer segment: stacks of membrane discs containing photopigment molecules
- Rods: discs are free-floating within the outer segment
- Cones: discs are continuous with the outer membrane
- Inner segment: contains mitochondria and ribosomes (metabolic machinery)
- Nucleus
- Synaptic terminal: ribbon synapse releasing glutamate onto bipolar and horizontal cells
Phototransduction - How Light Becomes Electrical Signal
This cascade occurs in rod outer segments (and analogously in cones):
In darkness (dark current):
- cGMP is high (maintained by guanylyl cyclase)
- High cGMP keeps CNG channels (cyclic nucleotide-gated cation channels) open
- Inward Na⁺ and Ca²⁺ flow ("dark current") → rod membrane is depolarized (~-40 mV)
- Depolarized rod continuously releases glutamate onto bipolar cells
When light hits the outer segment:
-
Photon absorbs into rhodopsin (or cone opsin) → 11-cis retinal isomerizes to all-trans retinal → rhodopsin becomes activated as metarhodopsin II (R*)
-
R* activates transducin (G-protein, Gt) → α-subunit dissociates → activates phosphodiesterase (PDE)
-
PDE degrades cGMP → cGMP levels fall
-
CNG channels close → Na⁺ and Ca²⁺ entry stops → hyperpolarization (membrane potential → ~-70 mV)
-
Ca²⁺ also falls (Na/Ca-K exchanger continues pumping Ca²⁺ out) → guanylyl cyclase is activated (recovery signal)
-
Reduced glutamate release from the photoreceptor terminal
Signal amplification: One photon → one R* → activates ~500 transducins → each activates one PDE → each PDE hydrolyzes ~1000 cGMP/second → massive amplification cascade. A single photon can be reliably detected.
Recovery of rhodopsin:
- Rhodopsin kinase phosphorylates R* → arrestin binds → quenches activation
- All-trans retinal dissociates → reduced to all-trans retinol → transported to RPE → re-isomerized to 11-cis retinal → returned to rod → regenerates rhodopsin (the visual cycle)
Dark adaptation: After exposure to bright light, rhodopsin bleaches. Recovery of sensitivity in the dark follows a time course: cone dark adaptation (~5-7 min); rod dark adaptation (~20-30 min, much slower because rod rhodopsin regeneration via RPE is slow).
Retinal Neural Processing
Light-induced changes in photoreceptor glutamate release activate the downstream retinal circuits:
Bipolar Cells:
- Two types responding to glutamate:
- ON-bipolar cells: have mGluR6 receptors (metabotropic) → glutamate causes hyperpolarization → IN DARK, constant glutamate hyperpolarizes them; IN LIGHT, reduced glutamate removes inhibition → they depolarize ("ON" response)
- OFF-bipolar cells: have ionotropic glutamate receptors (AMPA/KA) → glutamate depolarizes them; reduced glutamate in light → they hyperpolarize ("OFF" response)
Horizontal Cells:
- Connect photoreceptors laterally at the outer plexiform layer
- Mediate lateral inhibition → sharpen contrast (center-surround antagonism in the receptive field)
Amacrine Cells:
- ~30 different types; connect bipolar cells to ganglion cells at the inner plexiform layer
- Important for detecting motion, adjusting for background illumination (AII amacrine cells are key in the rod pathway)
Ganglion Cells:
- The only retinal cells that generate action potentials (all other retinal neurons use graded potentials)
- Their axons form the optic nerve (CN II)
- Have center-surround receptive fields:
- ON-center cells: depolarize to light in center of receptive field; inhibited by light in surround
- OFF-center cells: inhibited by light in center; depolarize to light in surround
- This organization maximizes sensitivity to contrast and edges
Major ganglion cell types:
- M cells (Magnocellular / Parasol cells): Large cells, fast-conducting, sensitive to movement and low contrast; color insensitive → project to Magnocellular layers of LGN
- P cells (Parvocellular / Midget cells): Small cells, slow, sensitive to fine detail and color; dominant in fovea → project to Parvocellular layers of LGN
- K cells (Koniocellular): Small bistratified cells; important for blue-yellow color → project to Koniocellular layers of LGN
- ipRGCs (intrinsically photosensitive RGCs): Contain melanopsin (opsin 4); respond directly to sustained bright light; project to the suprachiasmatic nucleus (circadian rhythm) and olivary pretectal nucleus (pupillary light reflex); NOT involved in image formation
CENTRAL VISUAL PATHWAYS: FROM RETINA TO VISUAL CORTEX
Overview of the Pathway
Retina → Optic nerve → Optic chiasm → Optic tract → Lateral Geniculate Nucleus (LGN) of thalamus → Optic radiations → Primary visual cortex (V1)
Plus important subcortical targets: superior colliculus, pretectal nuclei, suprachiasmatic nucleus
Step 1: Optic Nerve (CN II)
- Formed by the axons of ~1-1.2 million retinal ganglion cells
- Exits the globe at the optic disc → travels in the optic canal through the sphenoid bone
- Surrounds by meninges (dura, arachnoid, pia) and surrounded by CSF in the subarachnoid space (which is why raised ICP → papilledema)
- The nerve fibers maintain a retinotopic organization: superior retinal fibers run superior in the nerve, inferior fibers run inferior; macular fibers occupy a large central territory
Crucially, the retina represents a visual FIELD, not the whole world:
- The nasal retina sees the temporal visual field
- The temporal retina sees the nasal visual field
- The upper retina sees the inferior visual field
- The nasal half of each retina receives input from the same side's temporal visual hemifield
Step 2: Optic Chiasm
Located just above the pituitary gland (sella turcica) - clinically critical location.
At the chiasm, a partial decussation occurs:
- Nasal retinal fibers CROSS to the contralateral optic tract
- Temporal retinal fibers do NOT cross - they continue ipsilaterally
Consequence: Each optic tract carries information from the contralateral visual hemifield:
- Left optic tract = left temporal retina (right nasal visual field) + right nasal retina (right temporal visual field) → carries the entire right visual hemifield
- Right optic tract → carries the entire left visual hemifield
Clinical importance of the chiasm:
| Lesion location | Visual field defect |
|---|---|
| Optic nerve (before chiasm) | Monocular blindness (same eye) |
| Optic chiasm center | Bitemporal hemianopia (both temporal fields lost - classic pituitary tumor sign) |
| Lateral chiasm (both sides) | Binasal hemianopia (rare) |
| Optic tract | Contralateral homonymous hemianopia (incongruous) |
| Optic radiation (partial) | Quadrantanopia (see below) |
| Visual cortex | Contralateral homonymous hemianopia (with macular sparing) |
Step 3: Optic Tract
- Runs from the chiasm around the cerebral peduncles to the thalamus
- Contains fibers organized retinotopically:
- Superior retinal fibers (inferior visual field) run medially
- Inferior retinal fibers (superior visual field) run laterally
The optic tract terminates in three main places:
- Lateral geniculate nucleus (LGN) - ~90% of fibers → conscious vision
- Superior colliculus - reflexive eye movements (saccades, orienting)
- Pretectal nuclei - pupillary light reflex
- Suprachiasmatic nucleus (via melanopsin RGCs) - circadian rhythms
Step 4: Lateral Geniculate Nucleus (LGN)
The LGN is the thalamic relay for the visual pathway - a 6-layered structure in the pulvinar region of the dorsal thalamus.
Laminar organization:
- Layers 1 and 2 = Magnocellular (M) layers - receive input from M-type ganglion cells (movement, low contrast)
- Layers 3, 4, 5, 6 = Parvocellular (P) layers - receive input from P-type ganglion cells (detail, color)
- Between each layer: Koniocellular (K) sublayers - receive K ganglion cells (blue-yellow color)
Eye input segregation:
- Layers 1, 4, 6 receive input from the contralateral eye (nasal retinal fibers that crossed)
- Layers 2, 3, 5 receive input from the ipsilateral eye (temporal retinal fibers that did not cross)
- The two eyes are kept SEPARATE until V1
Retinotopy: The entire contralateral visual hemifield is mapped precisely onto each LGN. The fovea is disproportionately represented (magnification factor).
The LGN is NOT a simple relay - it receives massive feedback from V1 (corticogeniculate projections) and from the brainstem, which modulates signal transmission based on attention, arousal, and eye movements.
Step 5: Optic Radiations (Geniculocalcarine Tract)
The LGN projects to V1 via the optic radiations (also called the geniculocalcarine tract). This tract takes two distinct routes through the temporal and parietal lobes:
Superior fibers (parietal lobe route):
- Carry information from the inferior visual field (upper retina)
- Travel through the parietal white matter to the superior bank of the calcarine sulcus
Inferior fibers (Meyer's loop):
- Carry information from the superior visual field (lower retina)
- Loop anteriorly into the temporal lobe around the tip of the inferior horn of the lateral ventricle - this is Meyer's loop
- Then sweep posteriorly to reach the inferior bank of the calcarine sulcus
Clinical significance of Meyer's loop:
- Temporal lobe lesions (e.g., tumor, surgical resection for epilepsy) can damage Meyer's loop
- This causes contralateral superior quadrantanopia - "pie in the sky" defect (loss of upper contralateral visual field)
- Parietal lobe lesions damage the superior fibers → contralateral inferior quadrantanopia - "pie on the floor"
Step 6: Primary Visual Cortex (V1) - Striate Cortex
Location:
- Lies in and around the calcarine sulcus on the medial surface of the occipital lobe
- Also called Brodmann area 17, or the striate cortex (due to the white stripe of Gennari - myelinated fibers of layer IVb visible to the naked eye)
Retinotopic map:
- The entire contralateral visual hemifield is mapped onto V1
- Posterior pole of the occipital lobe: represents the fovea/central vision (large cortical area due to cortical magnification)
- Anterior calcarine area: represents the peripheral visual field
- Superior visual field → inferior bank of calcarine sulcus
- Inferior visual field → superior bank of calcarine sulcus
Cortical magnification factor: The fovea, though physically small, commands a disproportionately large area of V1 (~50-60% of V1 represents the central 10° of vision). This is called the cortical magnification factor and explains why central vision has so much higher resolution.
Layers of V1 and Parallel Processing:
V1 has 6 cortical layers with distinct inputs and outputs:
- Layer IV (especially IVCα, IVCβ): Receives direct LGN input
- IVCα: M-pathway input (magnocellular)
- IVCβ: P-pathway input (parvocellular)
- Layer II and III: Contain cytochrome oxidase-rich patches called blobs (color processing) and interblob regions (orientation selectivity)
- Layer V: Projects to superior colliculus
- Layer VI: Projects back to LGN (corticogeniculate feedback)
- Layer II/III interblobs and Layer IVB: Projects to extrastriate areas
Orientation selectivity (Hubel and Wiesel, Nobel Prize 1981):
- V1 neurons respond NOT to spots of light but to oriented bars or edges at specific angles
- Each neuron has a preferred orientation
- Neurons with similar orientation preferences are grouped in vertical columns: orientation columns
- Ocular dominance columns: adjacent vertical slabs responding preferentially to one eye or the other
- The full set of orientations and eye dominance preferences repeats across V1 as a mosaic - the hypercolumn concept
Step 7: Extrastriate Cortex - The Two Visual Streams
From V1, visual information is processed in a hierarchical fashion through extrastriate areas (V2, V3, V4, V5/MT, etc.) via two major parallel streams:
THE VENTRAL STREAM (the "What" Pathway):
- V1 → V2 → V4 → Inferior temporal (IT) cortex (areas TE, TEO)
- Processes: Form, color, object identity, face recognition, texture
- Damage → Agnosia (inability to recognize objects/faces despite intact primary vision)
- Color information (from P and K pathways) is important here
- Area V4: specialized for color constancy and wavelength processing
- Inferior temporal cortex: highest-level object recognition - neurons respond to complex shapes/faces regardless of size or position
THE DORSAL STREAM (the "Where/How" Pathway):
- V1 → V2 → V3 → V5/MT (middle temporal area) → Posterior parietal cortex (areas MT+, V3A, LIP, VIP)
- Processes: Motion, spatial location, depth, visuomotor guidance
- Damage to V5/MT → Akinetopsia (inability to perceive motion - the world looks like a series of still photographs)
- Damage to parietal cortex → Spatial neglect, optic ataxia (unable to reach accurately), Balint syndrome
- M-pathway dominates this stream
Subcortical Visual Pathways (Non-Geniculo-Striate)
1. Pupillary Light Reflex (PLR):
- Pathway: Retina → optic nerve → optic chiasm → optic tract → Pretectal nuclei (olivary pretectal nucleus) in midbrain → bilateral Edinger-Westphal nuclei → ciliary ganglion → sphincter pupillae → pupil constriction
- Both eyes constrict when one eye is illuminated: Direct reflex (same eye) and consensual reflex (other eye)
- Afferent limb: CN II (optic nerve); efferent limb: CN III (oculomotor, parasympathetic)
- Clinical use: RAPD (Relative Afferent Pupillary Defect / Marcus Gunn pupil) - afferent defect on one side → when swinging flashlight from good to bad eye, bad eye's pupil dilates paradoxically
2. Visual Component of the Accommodation Reflex:
- Retina → V1 → Frontal Eye Fields → midbrain → CN III → ciliary muscle (accommodation) + sphincter pupillae (miosis) + medial recti (convergence) = near triad
3. Superior Colliculus:
- Receives direct retinal input (~10% of optic tract fibers)
- Controls reflexive saccadic eye movements (orienting gaze toward novel stimuli)
- Integrated with the frontal eye fields and parietal cortex for voluntary saccades
4. Suprachiasmatic Nucleus (SCN) - Circadian Rhythm:
- Receives direct retinal input from melanopsin-containing ipRGCs
- Acts as the master circadian clock
- Entrains the body's ~24-hour rhythms to the environmental light/dark cycle
Summary: Complete Visual Signal Flow
Light → Retina (phototransduction: photon → rhodopsin → transducin → PDE → ↓cGMP → CNG closure → hyperpolarization → ↓glutamate release)
↓
Bipolar cells (ON/OFF channels split here)
↓
Retinal ganglion cells (action potentials; M/P/K streams; center-surround receptive fields)
↓
Optic nerve (CN II) → Optic chiasm (nasal fibers cross) → Optic tract
↓
LGN (6 layers; M/P/K streams; eye segregated; contralateral visual hemifield)
↓
Optic radiations → Meyer's loop (superior field) / parietal route (inferior field)
↓
V1 - Primary visual cortex (striate cortex, Brodmann 17, calcarine sulcus)
→ Orientation columns, ocular dominance columns, hypercolumns
↓
Extrastriate cortex:
VENTRAL stream → V4 → IT cortex: "WHAT" (color, form, object identity)
DORSAL stream → V5/MT → Parietal cortex: "WHERE/HOW" (motion, spatial)
Sources: Ganong's Review of Medical Physiology, 26th Edition (Chapters 10-11)
This is a shared conversation. Sign in to Orris to start your own chat.