Explain In Detail The Neural And Chemical Regulation Of Respiration******.
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
I now have comprehensive content and diagrams. Let me compile the full detailed answer.
Neural and Chemical Regulation of Respiration
Sources: Guyton and Hall Textbook of Medical Physiology (13th ed.) and Costanzo Physiology (7th ed.)
Overview
Respiration is automatically adjusted from birth until death to accommodate eating, talking, sleeping, exercise, and changing environments. The nervous system coordinates these adjustments to maintain arterial PO₂, PCO₂, and H⁺ within near-normal limits even during heavy exercise. The regulatory system has two major arms: neural control (structural organization of brain stem centers) and chemical control (responses to CO₂, H⁺, and O₂).
PART I: NEURAL REGULATION OF RESPIRATION
1. The Respiratory Center
The respiratory center is composed of several bilaterally distributed neuron groups in the medulla oblongata and pons. It is divided into three major collections:
- Dorsal Respiratory Group (DRG) - primarily controls inspiration
- Ventral Respiratory Group (VRG) - contains both inspiratory and expiratory neurons
- Pneumotaxic Center - controls rate and depth of breathing
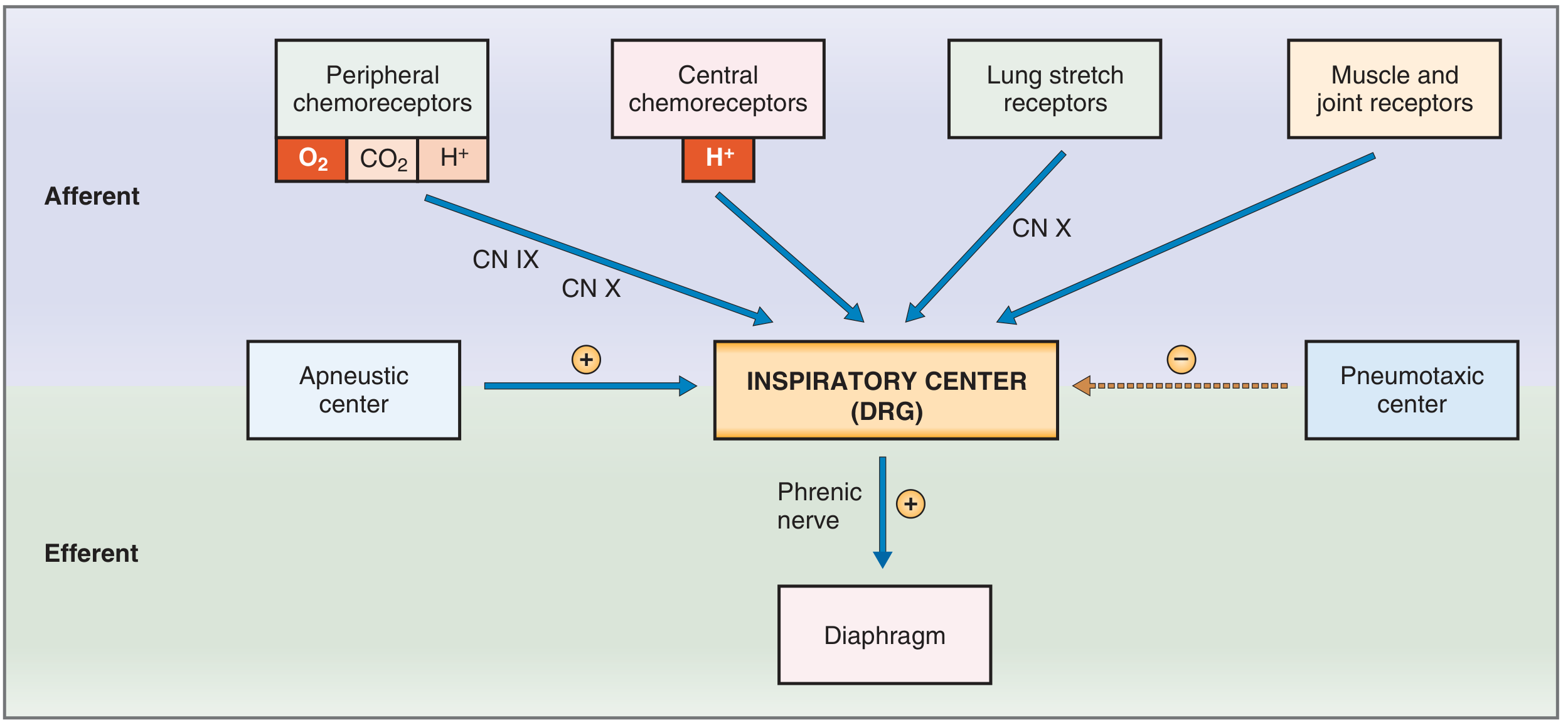
Fig. 5.32 - Costanzo Physiology: Brain stem control of breathing showing afferent and efferent pathways.
2. Dorsal Respiratory Group (DRG) - Controls Inspiration and Rhythm
The DRG extends most of the length of the medulla. Most of its neurons are located in the nucleus of the tractus solitarius (NTS), which is the sensory termination of the vagal and glossopharyngeal nerves. It receives sensory signals from:
- Peripheral chemoreceptors
- Baroreceptors
- Receptors in the liver, pancreas, and GI tract
- Several types of receptors in the lungs
Rhythmical Inspiratory Discharges: Even when all peripheral nerves entering the medulla are sectioned and the brain stem is transected above and below the medulla, the DRG still emits repetitive bursts of inspiratory neuronal action potentials. In humans, neural networks are present where one set of neurons excites a second set, which in turn inhibits the first - then after a period, the cycle repeats. This produces the basic respiratory rhythm.
Pre-Bötzinger Complex: A small region of the rostral VRG called the pre-Bötzinger complex contains spontaneously firing neurons that project to both the DRG and VRG, contributing importantly to the basic rhythm.
Ramp Signal for Inspiration: The DRG neurons send a progressively increasing ("ramp") signal to the inspiratory muscles. Inspiration begins slowly, builds over ~2 seconds as the lungs fill, then abruptly cuts off for about 3 seconds (expiration) - the cycle then repeats. The advantages of a ramp signal include: (1) gradual increase in lung volume during each breath rather than sudden inflation, and (2) ease of controlling the rate and depth of respiration by changing the rate of rise and the duration of the ramp.
3. Ventral Respiratory Group (VRG) - Inspiration and Expiration
The VRG is located in the ventrolateral medulla (nucleus ambiguus and nucleus retroambiguus). It contains both inspiratory and expiratory neurons. During quiet breathing, the VRG is almost entirely inactive - quiet expiration is a passive process from elastic recoil. During forceful breathing, signals spread into the VRG to provide extra drive for both inspiratory and expiratory muscles. The expiratory neurons drive the abdominal and other expiratory muscles when active expiration is needed (e.g., during exercise).
4. Pneumotaxic Center - Limits Duration of Inspiration
The pneumotaxic center is located in the nucleus parabrachialis in the dorsal upper pons. Its main function is to limit inspiration - it transmits signals to the inspiratory area that switch off the ramp signal, thereby limiting the duration of the ramp. When the pneumotaxic signal is strong, inspiration may last as little as 0.5 seconds, filling the lungs only slightly. When the signal is weak, inspiration may continue for 5 seconds or more. The net effect is to increase the respiratory rate when the signal is strong, because the shortened inspiration is followed by proportionally shortened expiration.
5. Apneustic Center - Prolongs Inspiration
Located in the lower pons, the apneustic center provides a tonic excitatory drive to the inspiratory center (DRG), prolonging the burst of action potentials in the phrenic nerve and prolonging contraction of the diaphragm. It is normally held in check by the pneumotaxic center. If the pneumotaxic center is eliminated and vagal inputs are severed, apneusis results - an abnormal pattern of prolonged inspiratory gasps followed by brief expiratory movements.
6. The Hering-Breuer Inflation Reflex
Stretch receptors in the bronchi and bronchioles transmit signals through the vagus nerves to the DRG. When the lungs become overly inflated, these receptors send signals that switch off inspiration and increase expiratory time - the Hering-Breuer reflex. In humans, this reflex is not activated until tidal volume increases to more than 3x normal (~1.5 L/breath). It functions mainly as a protective mechanism preventing excess lung inflation rather than as a major controller of normal ventilation.
7. Other Mechanoreceptors Influencing Respiration
| Receptor Type | Location | Effect |
|---|---|---|
| Lung stretch receptors | Airway smooth muscle | Hering-Breuer reflex: inhibits inspiration |
| Joint and muscle receptors | Joints and muscles of limbs | Stimulate inspiratory center; important in early exercise response |
| Irritant receptors | Airway epithelium | Stimulate coughing, bronchoconstriction |
| Juxtacapillary (J) receptors | Pulmonary capillaries | Stimulated by pulmonary edema/congestion; cause rapid shallow breathing |
8. Higher Center Influences
Cerebral cortex: Can temporarily override the automatic brain stem centers. A person can voluntarily hyperventilate (which decreases PaCO₂ and is self-limiting - eventually causing unconsciousness) or voluntarily hypoventilate (breath-holding). A period of prior hyperventilation prolongs breath-holding duration by lowering the PaCO₂ before the apneic period.
Hypothalamus: Contributes to respiratory adjustments during temperature changes, emotional states, and other autonomic responses.
PART II: CHEMICAL REGULATION OF RESPIRATION
The ultimate goal of respiration is to maintain proper concentrations of O₂, CO₂, and H⁺ in the tissues. Chemical control responds to changes in all three substances, but in very different ways.
Key principle: CO₂ and H⁺ act directly on the central respiratory center in the medulla. Oxygen acts almost entirely through peripheral chemoreceptors.
9. Central Chemoreceptors - CO₂ and H⁺
Location: The retrotrapezoid nucleus and other neurons in the ventrolateral medulla, lying only ~0.2 mm beneath the ventral surface of the rostral medulla.
Primary stimulus: H⁺ ions are the primary direct stimulus for these neurons. However, H⁺ does not easily cross the blood-brain barrier, so changes in blood H⁺ concentration have considerably less effect than changes in blood CO₂.
How CO₂ works indirectly:
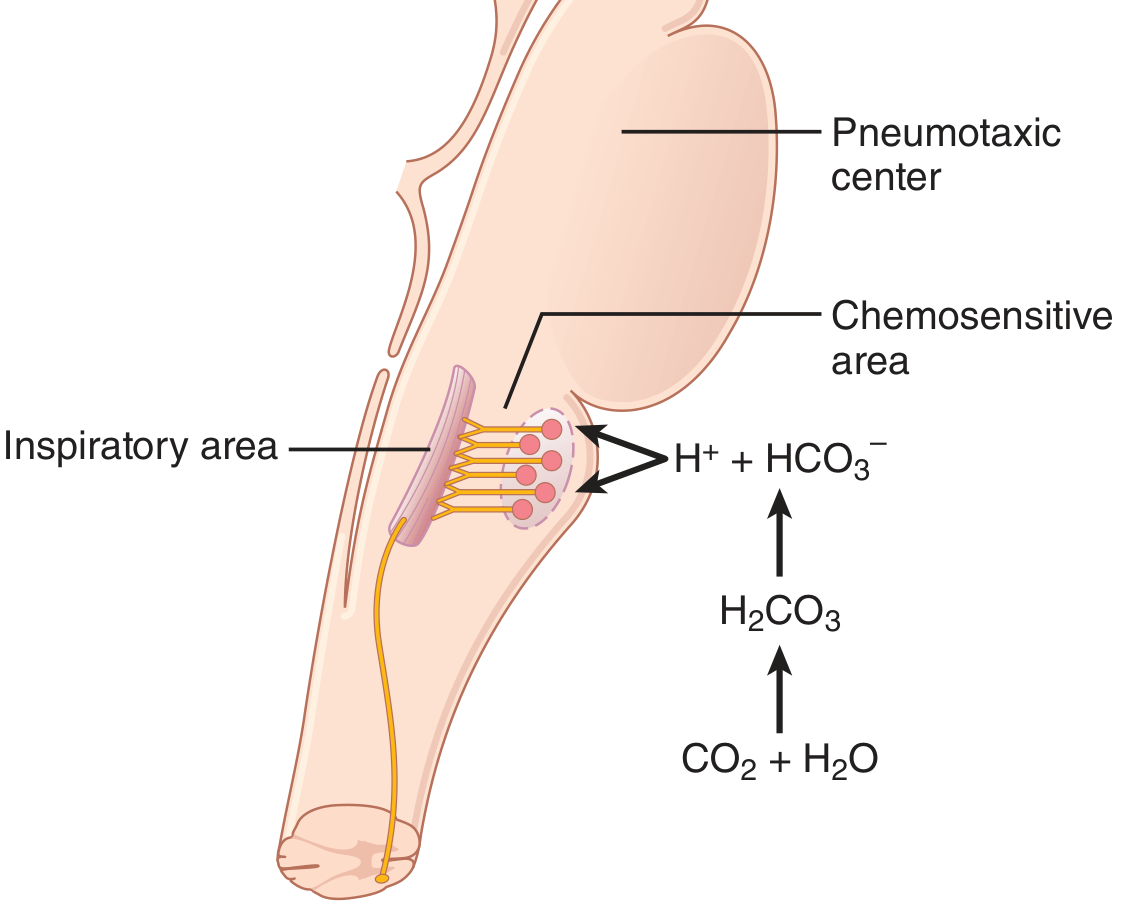
Figure 42.2 - Guyton & Hall: CO₂ stimulates the chemosensitive area indirectly by generating H⁺ after crossing the blood-brain barrier.
The steps are:
- In blood, CO₂ combines with H₂O to form H⁺ and HCO₃⁻ - but these ions cannot cross the blood-brain barrier
- CO₂ itself freely crosses the blood-brain barrier into brain ECF and CSF
- In the CSF, CO₂ reacts with water: CO₂ + H₂O → H₂CO₃ → H⁺ + HCO₃⁻
- The H⁺ generated in CSF directly stimulates the central chemoreceptors
- Central chemoreceptors signal the inspiratory center to increase breathing rate (hyperventilation)
Why CO₂ is more potent than blood H⁺: Because CO₂ crosses the blood-brain barrier freely while H⁺ and HCO₃⁻ are trapped in the blood, a rise in blood PCO₂ causes a disproportionately large rise in H⁺ in the CSF and brain ECF, generating a stronger respiratory stimulus than an equivalent rise in blood H⁺.
Quantitative effects: A rise in arterial PCO₂ in the range of 35-75 mmHg causes a marked increase in alveolar ventilation. By contrast, a change in blood pH in the range of 7.3-7.5 causes less than 10% as great a change in ventilation. This confirms that CO₂/PCO₂ is the dominant day-to-day chemical controller of respiration.
Adaptation over 1-2 days: The stimulatory effect of a sustained CO₂ rise declines over 1-2 days to about one-fifth the initial effect, because:
- The kidneys increase blood HCO₃⁻, which binds H⁺ in blood and CSF
- HCO₃⁻ slowly diffuses across the blood-brain barrier and reduces H⁺ near respiratory neurons
This explains why CO₂ changes have a potent acute effect but only a weak chronic effect on respiratory drive.
10. Peripheral Chemoreceptors - The Role of Oxygen
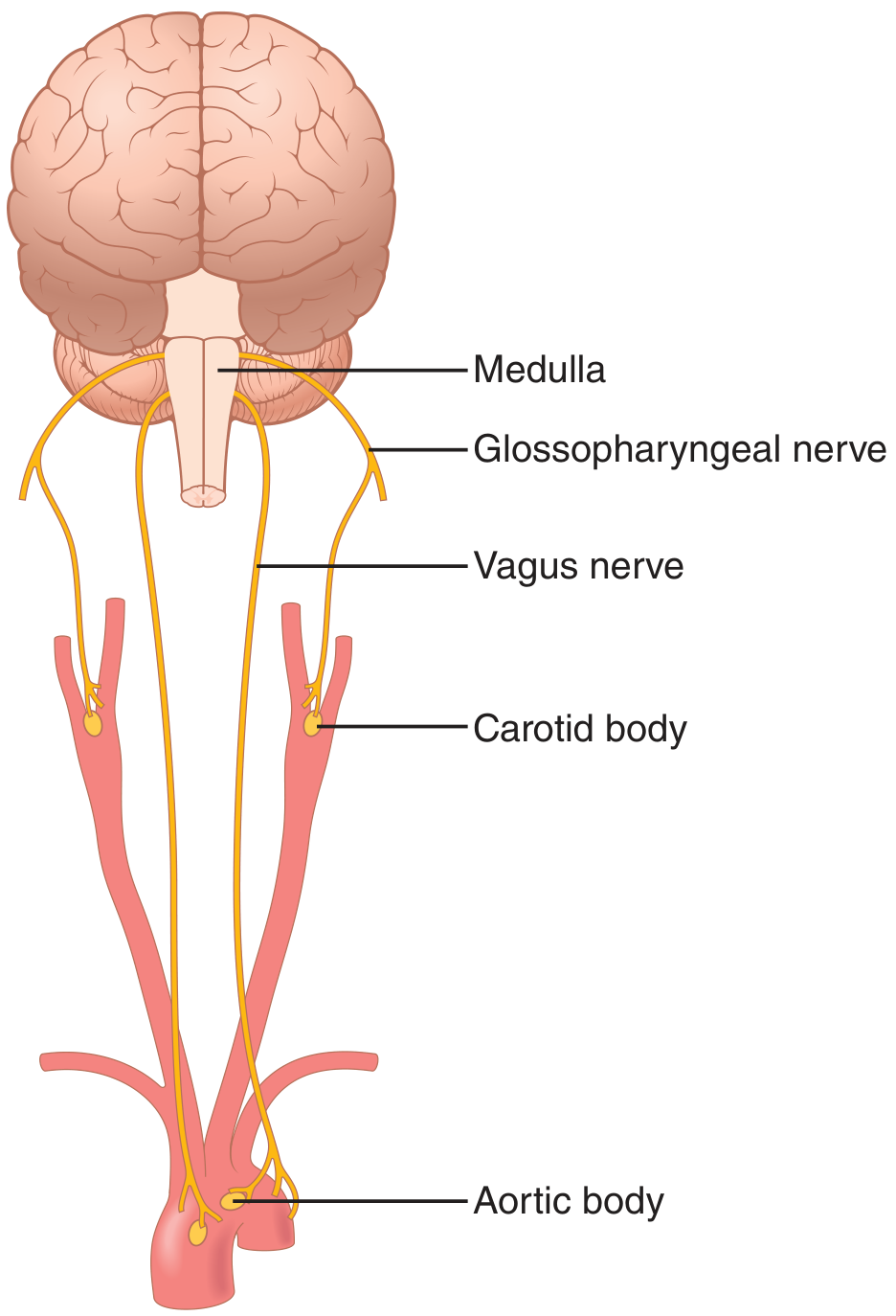
Figure 42.4 - Guyton & Hall: Peripheral chemoreceptors in the carotid and aortic bodies.
Locations:
- Carotid bodies - bilaterally at the bifurcation of the common carotid arteries; afferent fibers travel via Hering's nerves → glossopharyngeal nerve (CN IX) → DRG
- Aortic bodies - along the arch of the aorta; afferent fibers travel via the vagus nerves (CN X) → DRG
- A few additional chemoreceptors are associated with other thoracic and abdominal arteries
Blood supply: Each chemoreceptor body receives its own blood supply directly from the adjacent artery. Blood flow is extremely high - 20 times the weight of the body each minute - so essentially zero O₂ is extracted. This means the chemoreceptors are always exposed to arterial (not venous) blood.
Response to Decreased PO₂ (Hypoxic Drive)
The peripheral chemoreceptors are the body's O₂ sensors. Key features:
- Relatively insensitive to PO₂ changes between 100 and 60 mmHg - breathing rate is virtually constant in this range
- When arterial PO₂ falls below 60 mmHg, chemoreceptor firing rate increases steeply and linearly
- So sensitive below 60 mmHg that firing rate may change during a single breathing cycle
- At 20 mmHg, ventilation increases approximately 6-fold over normal
This threshold at 60 mmHg corresponds to the steep portion of the oxyhemoglobin dissociation curve, providing a clinically important safety margin.
Response to Increased PCO₂ and Decreased pH
- Peripheral chemoreceptors also respond to increased PCO₂, but this effect is less important than the central chemoreceptor response
- They respond to decreased arterial pH (H⁺ increase) - important in metabolic acidosis where they mediate the compensatory hyperventilation (Kussmaul breathing)
- Notably, the response to decreased pH is mediated only by carotid bodies, not aortic bodies
Comparison: Central vs. Peripheral Chemoreceptors
| Feature | Central Chemoreceptors | Peripheral Chemoreceptors |
|---|---|---|
| Location | Ventrolateral medulla (retrotrapezoid nucleus) | Carotid bodies, aortic bodies |
| Primary stimulus | H⁺ (from CO₂ hydration in CSF) | O₂ (also CO₂ and H⁺) |
| Most important function | Minute-to-minute CO₂/pH regulation | Detection of hypoxia (PO₂ <60 mmHg) |
| Response threshold | Sensitive across normal PCO₂ range | Dramatic response only below PO₂ 60 mmHg |
| Blood-brain barrier impact | H⁺ is trapped; CO₂ crosses freely | Directly bathed in arterial blood |
| Afferent pathway | Direct neural connection to DRG | CN IX / CN X → DRG |
11. Why O₂ Is Not the Primary Controller
The hemoglobin-O₂ buffer system delivers near-normal amounts of O₂ to tissues even when pulmonary PO₂ changes from as low as 60 mmHg up to 1000 mmHg. Adequate O₂ delivery can therefore occur despite ventilation ranging from slightly below half-normal to 20 or more times normal. This is not true for CO₂ - blood and tissue PCO₂ change inversely with the rate of pulmonary ventilation. Animal evolution has therefore made CO₂ the major controller of respiration, with O₂ serving as a backup emergency system for when tissues are genuinely in danger.
12. Composite Chemical Control
PCO₂, pH, and PO₂ interact in a complex but predictable way:
- At a given PO₂, increasing PCO₂ causes a steep rise in alveolar ventilation
- The same PCO₂ curve is shifted leftward (greater ventilation for same PCO₂) when PO₂ is low (40 vs. 100 mmHg)
- A decrease in blood pH to 7.3 shifts the entire family of curves leftward (i.e., acidosis potentiates the ventilatory response to CO₂)
- All three stimuli - hypercapnia, hypoxia, and acidosis - synergize to drive ventilation
PART III: REGULATION DURING EXERCISE
During strenuous exercise, O₂ consumption and CO₂ formation can increase 20-fold, yet alveolar ventilation increases almost exactly in step and arterial blood gases remain near-normal. How?
Neurogenic (anticipatory) mechanism: At the moment exercise begins, the brain transmits collateral nerve impulses from the motor cortex into the brain stem respiratory center simultaneously with signals going to the exercising muscles. This is analogous to simultaneous cardiovascular activation at exercise onset. Ventilation jumps immediately - before blood chemical changes have had time to occur, and so strongly that it initially decreases arterial PCO₂ slightly below normal.
Chemical fine-tuning: After 30-40 seconds, CO₂ released from active muscles matches the elevated ventilation rate and arterial PCO₂ returns to normal. From this point, chemical signals (CO₂, H⁺, lactic acid during intense exercise) fine-tune the ventilation to match metabolic demands precisely.
Joint and muscle proprioceptors: Mechanoreceptors in the joints and muscles also send early ("anticipatory") signals to the inspiratory center - important in the rapid rise in ventilation at exercise onset even before blood gas changes are detected.
Summary of exercise control:
- Onset: Neurogenic drive from motor cortex collaterals + joint/muscle proprioceptors → immediate ventilation increase
- Sustained: CO₂ and H⁺ build-up provide chemical feedback → maintain ventilation matched to metabolic rate
- Intense exercise: Lactic acidosis provides additional H⁺ stimulus via peripheral chemoreceptors
PART IV: ACCLIMATIZATION TO HIGH ALTITUDE
Chronic low O₂ (as at high altitude) produces a phenomenon called acclimatization:
- Over 2-3 days, the brain stem loses ~80% of its sensitivity to PCO₂ and H⁺
- Excess ventilation that would normally blow off CO₂ and inhibit respiration fails to fully occur
- Low O₂ can therefore drive ventilation to much higher levels than under acute conditions
- Acute hypoxia may produce ~70% increase in ventilation; after 2-3 days of acclimatization, alveolar ventilation can increase 400-500% above baseline
- This greatly improves O₂ delivery to tissues at altitude
Summary Flow Diagram
RESPIRATORY REGULATION
│
├── NEURAL (Brain Stem)
│ ├── Medulla
│ │ ├── DRG (NTS) → Inspiration, Basic Rhythm (via Pre-Bötzinger complex)
│ │ └── VRG → Active expiration (quiet expiration = passive)
│ └── Pons
│ ├── Pneumotaxic center (upper) → Inhibits inspiration → ↑Rate
│ └── Apneustic center (lower) → Excites inspiration → ↑Depth
│
├── CHEMICAL - Central (Medullary Chemoreceptors)
│ └── ↑PCO₂ → CO₂ crosses BBB → ↑CSF H⁺ → stimulates DRG → Hyperventilation
│ (most important minute-to-minute controller)
│
├── CHEMICAL - Peripheral (Carotid/Aortic Bodies)
│ ├── ↓PO₂ below 60 mmHg → ↑Ventilation (hypoxic drive)
│ ├── ↑PCO₂ (less important than central)
│ └── ↓pH (especially metabolic acidosis) → via carotid bodies only
│
└── HIGHER CENTERS
├── Cerebral cortex → Voluntary override
├── Hypothalamus → Autonomic/temperature responses
└── Motor cortex → Collateral signals during exercise
Sources:
- Guyton and Hall Textbook of Medical Physiology, 14th ed., Chapter 42 (pp. 531-538)
- Costanzo Physiology, 7th ed., Chapter 5 (pp. 239-241)
This is a shared conversation. Sign in to Orris to start your own chat.