Mutarotation of glucose short explanation and diagram
Reading File
Finding Sources
Searching the Web
mutarotation of glucose biochemistry mechanism alpha beta anomers diagram
Reading File
Reading File
Loading Image
Loading Image
Here is a concise explanation with both diagrams:
Mutarotation of Glucose
Mutarotation is the spontaneous interconversion between the α and β anomeric forms of glucose (and other sugars) in aqueous solution, passing through a transient open-chain (acyclic) intermediate. The word literally means "changing rotation" - it was first observed as a gradual shift in the optical rotation of a freshly dissolved sugar solution.
How it happens - step by step
-
Cyclization creates an anomeric carbon. In solution, the open-chain aldehyde of glucose reacts with the C-5 hydroxyl group to form a 6-membered (pyranose) ring. C-1 becomes the anomeric carbon - a new chiral centre.
-
Two anomers are created. Depending on the orientation of the new -OH at C-1:
- α-D-Glucopyranose - the C-1 -OH is axial (below the ring plane in Haworth; same side as the ring in Fischer projection). Specific rotation = +112°
- β-D-Glucopyranose - the C-1 -OH is equatorial (above the ring plane; opposite side to CH₂OH). Specific rotation = +18.7°
-
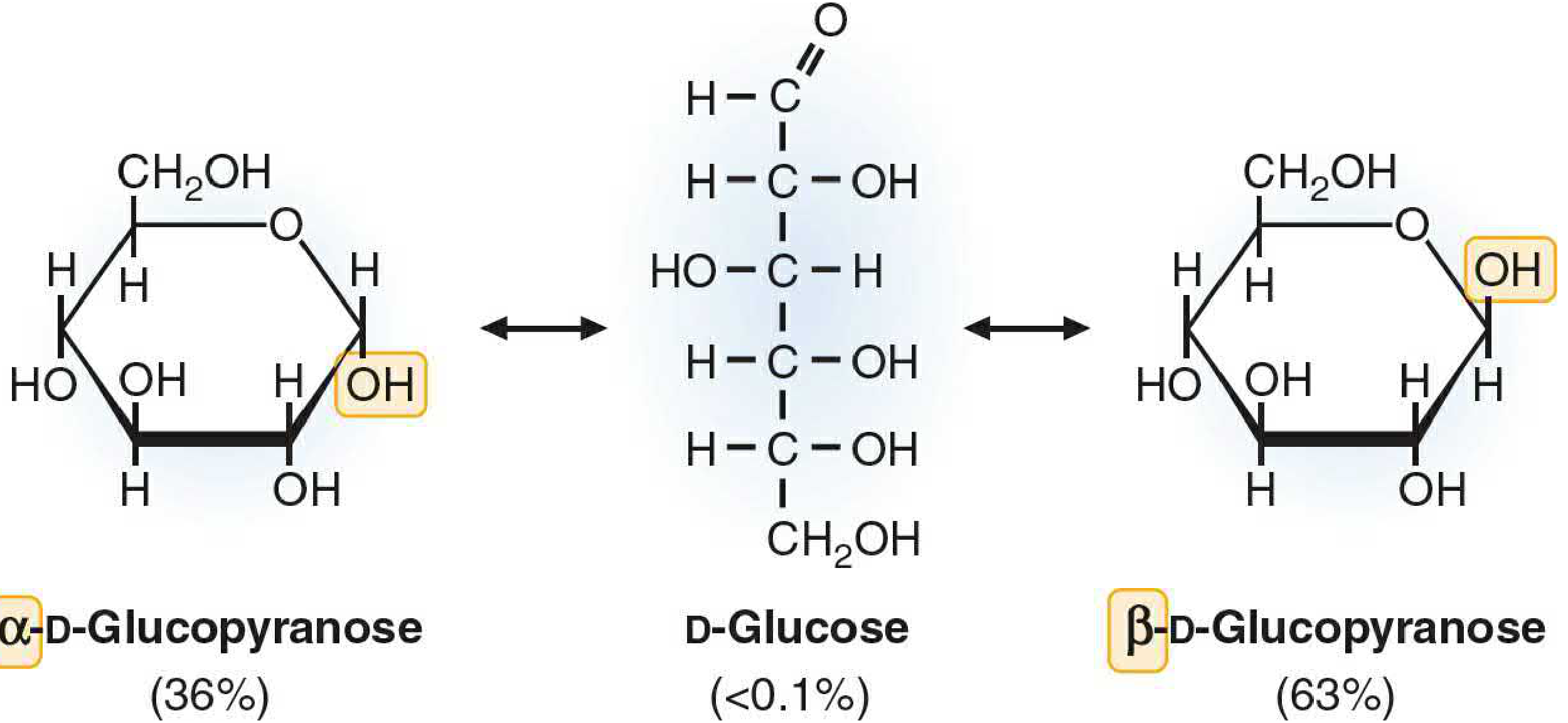
The ring opens and re-closes. Either anomer can open back to the linear aldehyde form (< 0.1% of molecules at any time). When the ring re-closes, C-1 can adopt either orientation - so the anomers slowly interconvert.
-
Equilibrium is reached. The optical rotation drifts until it settles at a stable value of +52.5°, corresponding to a mixture of:
- α form: ~36%
- β form: ~63-64%
- Open chain: < 0.1%
The β anomer dominates because the equatorial -OH position is sterically more stable.
Diagrams
Figure 1 - Summary diagram (optical rotation perspective):
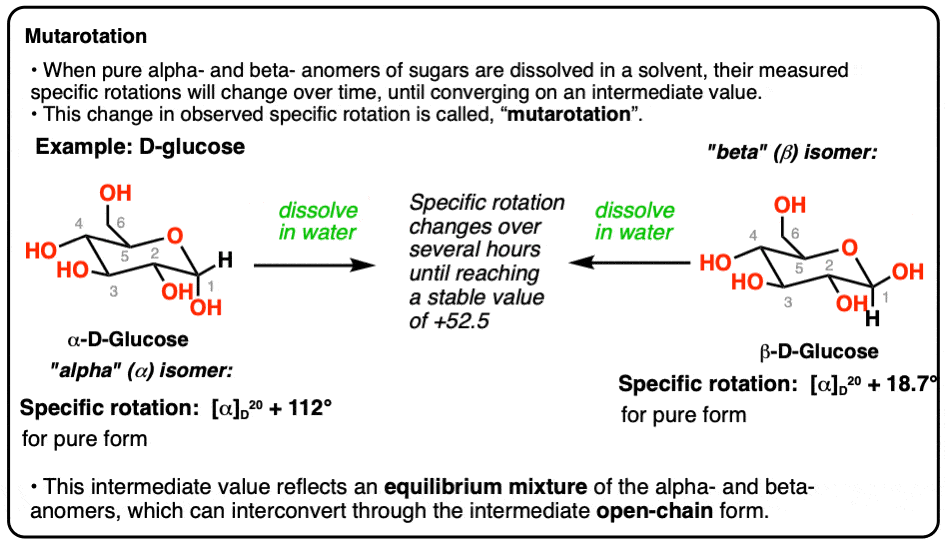
Figure 2 - Haworth projection showing the open-chain intermediate and equilibrium percentages (from Basic Medical Biochemistry - A Clinical Approach, 6e, Fig. 5.13):
Key points to remember
| Feature | α-D-Glucose | β-D-Glucose |
|---|---|---|
| C-1 -OH position | Axial (down) | Equatorial (up) |
| Specific rotation | +112° | +18.7° |
| Equilibrium proportion | 36% | 63% |
| Equilibrium [α] | +52.5° (shared) | +52.5° (shared) |
- Mutarotation is spontaneous but slow at neutral pH in pure water; it is accelerated by acid, base, or the enzyme mutarotase (aldose-1-epimerase) found in cells.
- If the anomeric carbon forms a glycosidic bond (e.g., in starch, cellulose, glycoproteins), mutarotation is blocked - the bond is locked in α or β configuration. This is why glycogen (α-1,4 bonds) and cellulose (β-1,4 bonds) have completely different biological properties.
- Clinically relevant in glucose oxidase assays: the enzyme is specific for β-D-glucose, so some commercial preparations include mutarotase to ensure complete conversion of α to β form for accurate blood glucose measurement.
Sources: Basic Medical Biochemistry - A Clinical Approach, 6e, p. 124; Biochemistry 8th ed. Lippincott Illustrated Reviews, p. 259; Tietz Textbook of Laboratory Medicine, 7th ed.
Invert sugar
Finding Sources
Searching the Web
invert sugar biochemistry sucrose hydrolysis invertase optical rotation
Reading File
Invert Sugar
Invert sugar is the equimolar (1:1) mixture of D-glucose and D-fructose produced by the hydrolysis of sucrose.
The Reaction
Sucrose (a disaccharide) is hydrolysed at its glycosidic bond - specifically the α,β-1,2-glycosidic bond linking glucose (C-1) to fructose (C-2):
Sucrose + H₂O → D-Glucose + D-Fructose
(C₁₂H₂₂O₁₁) (C₆H₁₂O₆) (C₆H₁₂O₆)
This is catalysed by:
- Invertase (β-fructofuranosidase, EC 3.2.1.26) - the enzyme in yeast, bees, and the intestinal brush border
- Sucrase (in intestinal mucosa) - same reaction
- Dilute acid (acid hydrolysis) - used industrially
Why is it called "Invert" Sugar?
The name refers to the inversion of optical rotation (measured by polarimetry):
| Sugar | Optical Rotation | Type |
|---|---|---|
| Sucrose | +66.5° | Dextrorotatory |
| D-Glucose | +52.5° | Dextrorotatory |
| D-Fructose | -92° | Strongly levorotatory |
| Invert sugar (mixture) | -20° | Levorotatory |
Fructose has such a strong levorotatory effect (-92°) that it overwhelms the dextrorotatory contribution of glucose (+52.5°). The net rotation of the product mixture is negative, which is the opposite (i.e., inverted) sign of the starting sucrose. This is why the process is called "inversion" and the product "invert sugar." (Harper's Illustrated Biochemistry, 32nd ed., p. 163)
Sucrose Structure - why hydrolysis releases both anomers
Sucrose is unique among common disaccharides: it is a non-reducing sugar because both anomeric carbons (C-1 of glucose and C-2 of fructose) are involved in the glycosidic bond, so neither is free. On hydrolysis, both anomeric -OH groups are released, and both products (glucose and fructose) undergo mutarotation in solution - contributing to the final mixed optical rotation value of -20°.
Biological and Clinical Relevance
- Digestion: Sucrase-isomaltase in the intestinal brush border hydrolyses dietary sucrose to invert sugar for absorption.
- Honey: Bees secrete invertase in their salivary glands; most of the sucrose in nectar is converted to invert sugar during honey making - this is why honey is primarily glucose + fructose.
- Food industry: Invert sugar syrup is sweeter than sucrose (fructose is ~1.7× sweeter than glucose), resists crystallisation, and retains moisture - making it valuable in confectionery, jams, and soft-centred chocolates.
- Sucrase deficiency: Congenital sucrase-isomaltase deficiency causes osmotic diarrhoea and bloating on sucrose ingestion in infants, due to unabsorbed sucrose drawing water into the gut.
Quick Summary
Sucrose (dextrorotatory, +66.5°) is hydrolysed by invertase into an equal mix of glucose and fructose. Because fructose is strongly levorotatory (-92°), the product mixture is levorotatory (-20°) - the rotation has been "inverted." This mixture is called invert sugar.
Coris cycle
Finding Sources
Searching the Web
Cori cycle biochemistry muscle liver lactate gluconeogenesis diagram
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Cori Cycle (Lactic Acid Cycle)
Named after Carl Ferdinand Cori and Gerty Cori (Nobel Prize, 1947), the Cori cycle is a metabolic cooperation between skeletal muscle (or RBCs) and the liver that recycles lactate back into glucose.
The Two-Organ Cycle
In Muscle / RBCs (anaerobic conditions):
- Glucose → 2 Pyruvate (glycolysis) → 2 Lactate (via LDH, regenerating NAD⁺)
- Net yield: 2 ATP per glucose
- Lactate exits into the bloodstream
In Liver:
- Lactate enters → oxidised back to Pyruvate (via LDH)
- Pyruvate → Glucose via gluconeogenesis
- Glucose released back into blood
- Net cost: 6 ATP per glucose synthesised
Diagrams
Figure 1 - Intertissue flow (Lippincott, Biochemistry 8th ed., Fig. 10.2):
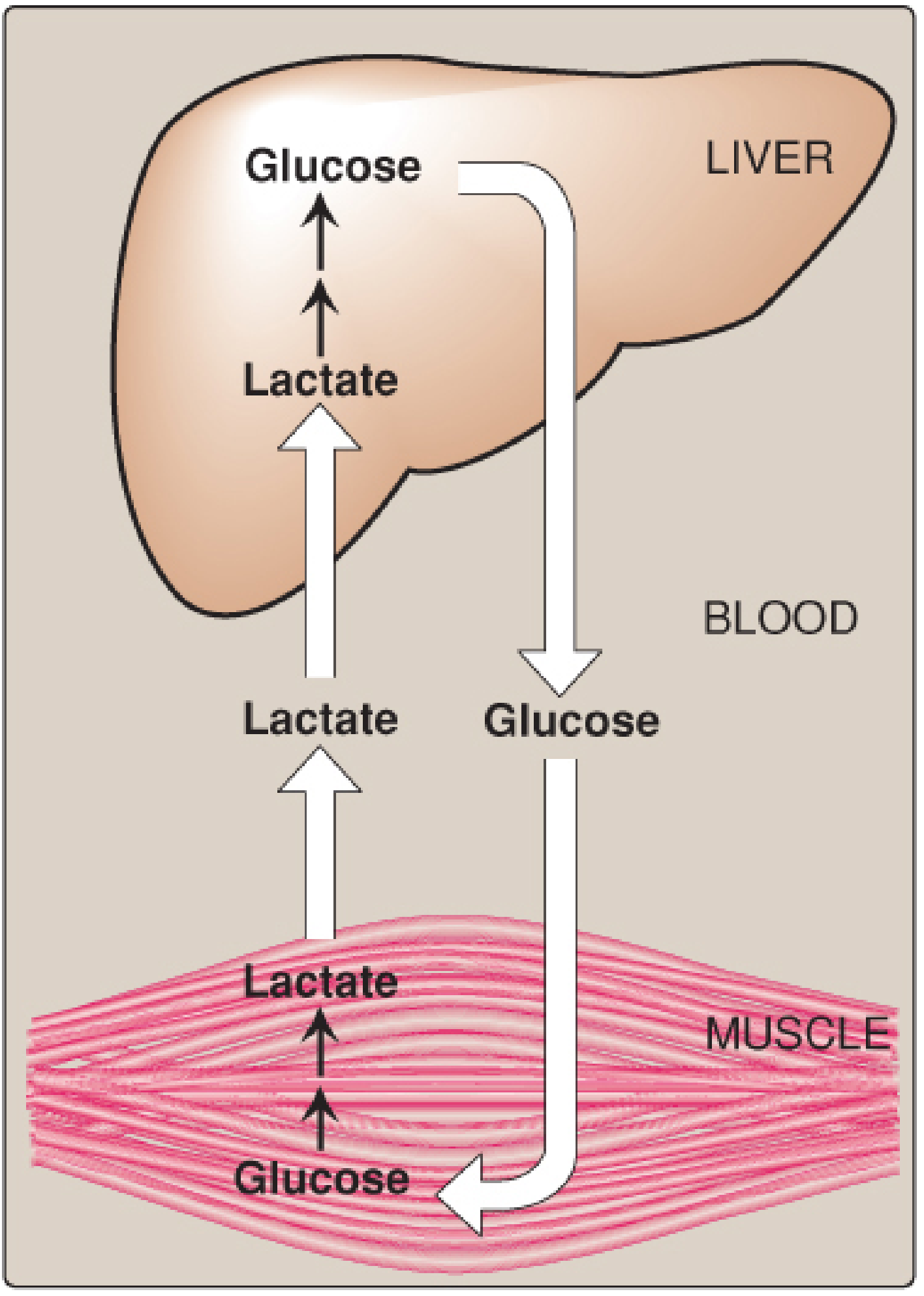
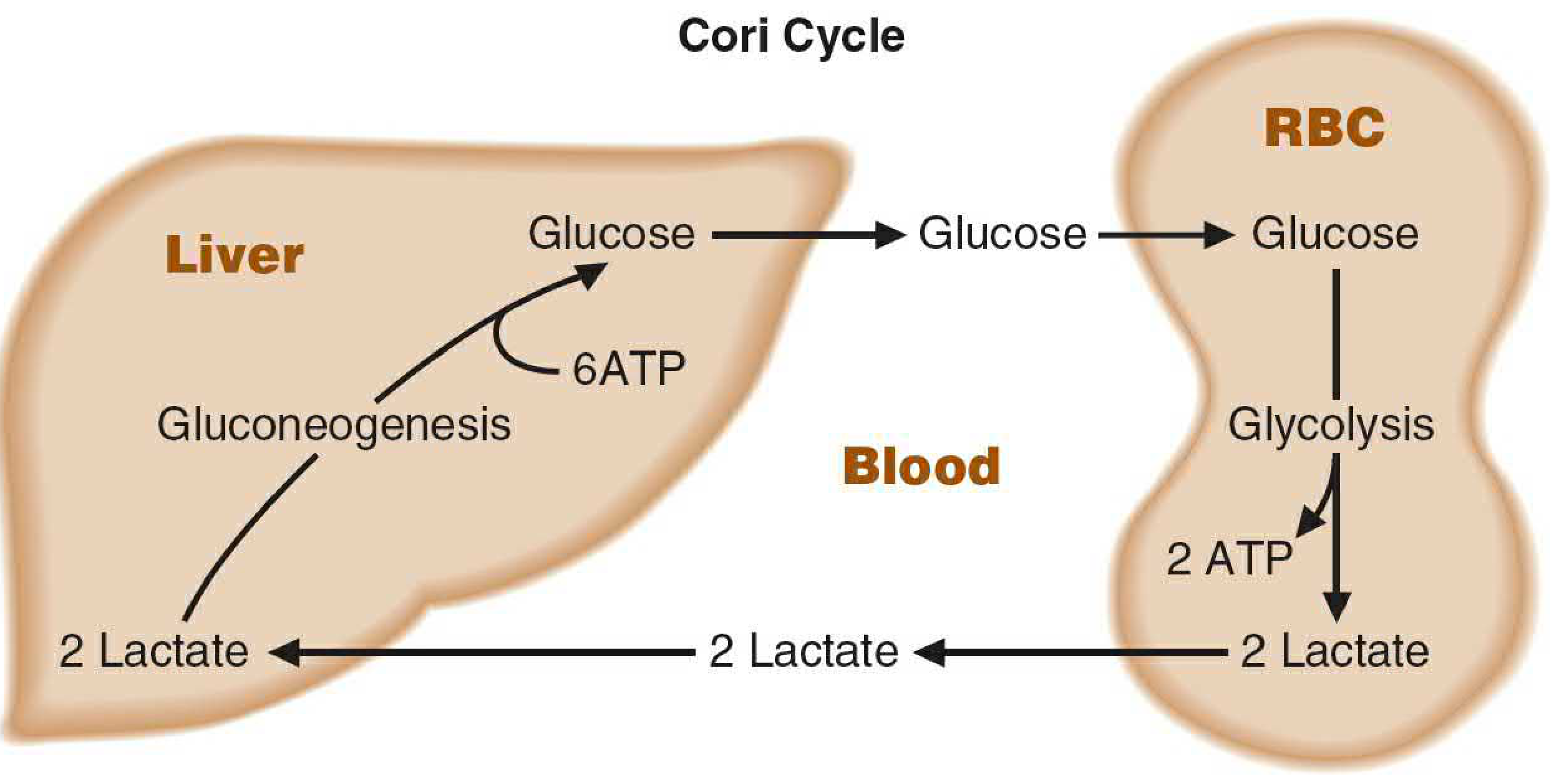
Figure 2 - ATP accounting across tissues (Basic Medical Biochemistry 6e, Fig. 22.12):
ATP Energy Balance - the "Metabolic Debt"
| Location | Process | ATP |
|---|---|---|
| Muscle / RBC | Glycolysis: Glucose → 2 Lactate | +2 ATP |
| Liver | Gluconeogenesis: 2 Lactate → Glucose | -6 ATP |
| Net | -4 ATP per cycle |
The cycle is energetically costly overall. The muscle "borrows" by running glycolysis cheaply; the liver "repays" the carbon debt at a higher energy cost. The liver's ATP comes from oxidative phosphorylation (fatty acid oxidation during fasting/exercise), so the net burden is shifted to a tissue that can sustain aerobic metabolism. (Biochemistry 8th ed. Lippincott, p. 355; Basic Medical Biochemistry 6e, p. 805)
Tissues Involved
- Lactate producers: Exercising skeletal muscle, red blood cells (no mitochondria so always anaerobic), white blood cells, intestinal mucosa, skin, brain astrocytes
- Lactate consumers (gluconeogenesis): Liver (primary), kidney cortex (especially during prolonged fasting)
- Other lactate consumers (oxidation): Heart muscle, resting skeletal muscle, brain neurons (fed by astrocytes)
Physiological Significance
- Prevents lactic acidosis during intense exercise - lactate is constantly cleared by the liver rather than accumulating.
- Maintains blood glucose during exercise and fasting - lactate is the second most important gluconeogenic substrate after amino acids.
- Recycles carbon skeletons - no carbon is lost; the same 6-carbon glucose backbone shuttles back and forth.
- Allows tissues without mitochondria (RBCs) to function - they are obligate glycolysers and depend on the liver to regenerate glucose from their lactate output.
Related Cycle - the Glucose-Alanine Cycle
A parallel cycle operates with alanine as the carbon carrier instead of lactate. In muscle, pyruvate is transaminated to alanine (simultaneously offloading NH₃), alanine travels to liver, and is converted back to pyruvate → glucose. This is sometimes called the Cahill cycle and is especially important during prolonged fasting.
"The cycling of lactate and glucose between peripheral tissues and liver is called the Cori cycle."
- Basic Medical Biochemistry: A Clinical Approach, 6e, p. 805
Epimer
Finding Sources
Searching the Web
epimers biochemistry glucose galactose mannose C2 C4 diagram
Reading File
Loading Image
Loading Image
Epimers
Definition: Epimers are carbohydrate isomers that differ in configuration at only one specific carbon atom, with the exception of the anomeric (carbonyl) carbon. They are a subtype of diastereomers - not mirror images, not identical, but differing at a single chiral centre. (Biochemistry 8th ed. Lippincott, p. 257)
Diagrams
Figure 1 - Fischer projection showing C-2 and C-4 epimers of glucose (Lippincott, Fig. 7.4):

Figure 2 - C-2 and C-4 epimers side by side:

The Classic Examples
| Pair | Differ at | Relationship |
|---|---|---|
| Glucose & Galactose | C-4 | C-4 epimers |
| Glucose & Mannose | C-2 | C-2 epimers |
| Galactose & Mannose | C-2 and C-4 | Isomers (NOT epimers) |
| Glucose & Fructose | Different carbonyl group | Isomers (NOT epimers) |
All four (glucose, galactose, mannose, fructose) share the same molecular formula C₆H₁₂O₆ - they are all hexoses and structural isomers of each other. But only pairs differing at a single carbon qualify as epimers.
How to Spot an Epimer (Fischer Projection)
Looking at the Fischer projection of D-Glucose as the reference:
C-1 CHO
C-2 H — OH ← swap here → Mannose (C-2 epimer)
C-3 HO — H
C-4 H — OH ← swap here → Galactose (C-4 epimer)
C-5 H — OH
C-6 CH₂OH
Swapping the -OH and -H at one carbon only = epimer. Swapping at two or more carbons = isomer (no longer an epimer).
Epimers vs Anomers - Important Distinction
| Feature | Epimer | Anomer |
|---|---|---|
| Carbon involved | Any chiral carbon except C-1 | Only C-1 (anomeric carbon) |
| How formed | Different compounds in solution | Ring opening/closing (mutarotation) |
| Examples | Glucose/Galactose (C-4), Glucose/Mannose (C-2) | α-glucose / β-glucose |
| Interconvert spontaneously? | No (need enzyme - epimerase) | Yes (mutarotation) |
Anomers are technically a special case of epimers (differing at one carbon), but by convention the term "epimer" is reserved for differences at carbons other than the anomeric carbon.
Biological Significance
- Galactose (C-4 epimer of glucose) must be converted to glucose-1-phosphate via the Leloir pathway (involving UDP-galactose 4-epimerase) before it can enter glycolysis. Failure of this conversion causes galactosaemia.
- Mannose (C-2 epimer of glucose) is converted to mannose-6-phosphate → fructose-6-phosphate (via phosphomannose isomerase) to enter glycolysis. Mannose is important in N-linked glycoprotein synthesis.
- Epimerases are the enzymes that interconvert epimers by inverting the -OH configuration at a specific carbon (e.g., UDP-galactose 4-epimerase, phosphomannose isomerase).
"Carbohydrate isomers that differ in configuration around only one specific carbon atom (with the exception of the carbonyl carbon) are defined as epimers of each other."
- Biochemistry 8th ed. Lippincott Illustrated Reviews, p. 257
This is a shared conversation. Sign in to Orris to start your own chat.