Why ketone bodies are used in starvation in muscle, brain, and heart?
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
I now have all the information needed for a thorough, well-illustrated answer. Here it is:
Ketone Bodies in Starvation: Why Muscle, Brain, and Heart Use Them
Background: What Are Ketone Bodies?
Ketone bodies are three water-soluble molecules produced by the liver from acetyl-CoA derived primarily from fatty acid beta-oxidation:
- Acetoacetate (the primary one)
- 3-Hydroxybutyrate (beta-hydroxybutyrate) - the most abundant in blood during starvation
- Acetone - a metabolic dead end (exhaled, not used for energy)
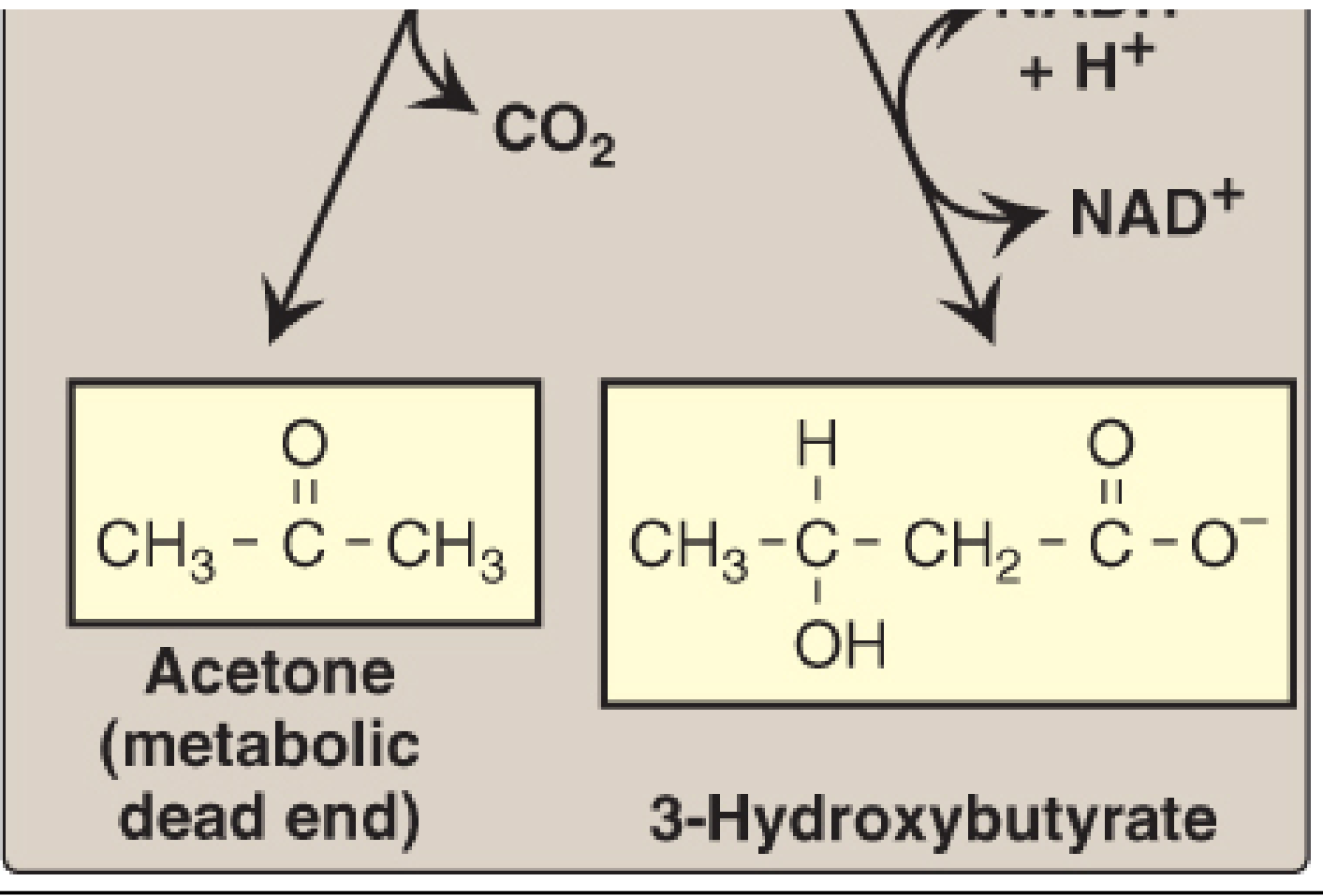
Why Ketone Bodies Rise During Starvation
During the first 3-7 days of fasting, the drop in insulin and rise in glucagon trigger several changes:
- Adipose tissue lipolysis accelerates, releasing free fatty acids (FFAs) into blood
- The liver takes up FFAs and runs beta-oxidation at a rate exceeding its own energy needs
- Excess acetyl-CoA in the liver is channeled into ketogenesis via the HMG-CoA pathway
- The liver peaks at ~100 g/day of ketone body production by day 3 of a fast
Plasma ketone bodies rise from negligible in the fed state to ~2.9 mmol/L at 40 hours and ~4.5 mmol/L after 7 days starvation, as shown in [Harper's Illustrated Biochemistry, 32nd Ed](book reference), while glucose only falls modestly (5.5 → 3.5 mmol/L).
How Peripheral Tissues Use Ketone Bodies (Ketolysis)
The liver produces but cannot use ketone bodies - it lacks thiophorase (succinyl-CoA:acetoacetate CoA transferase). All other mitochondria-containing tissues use the following pathway:
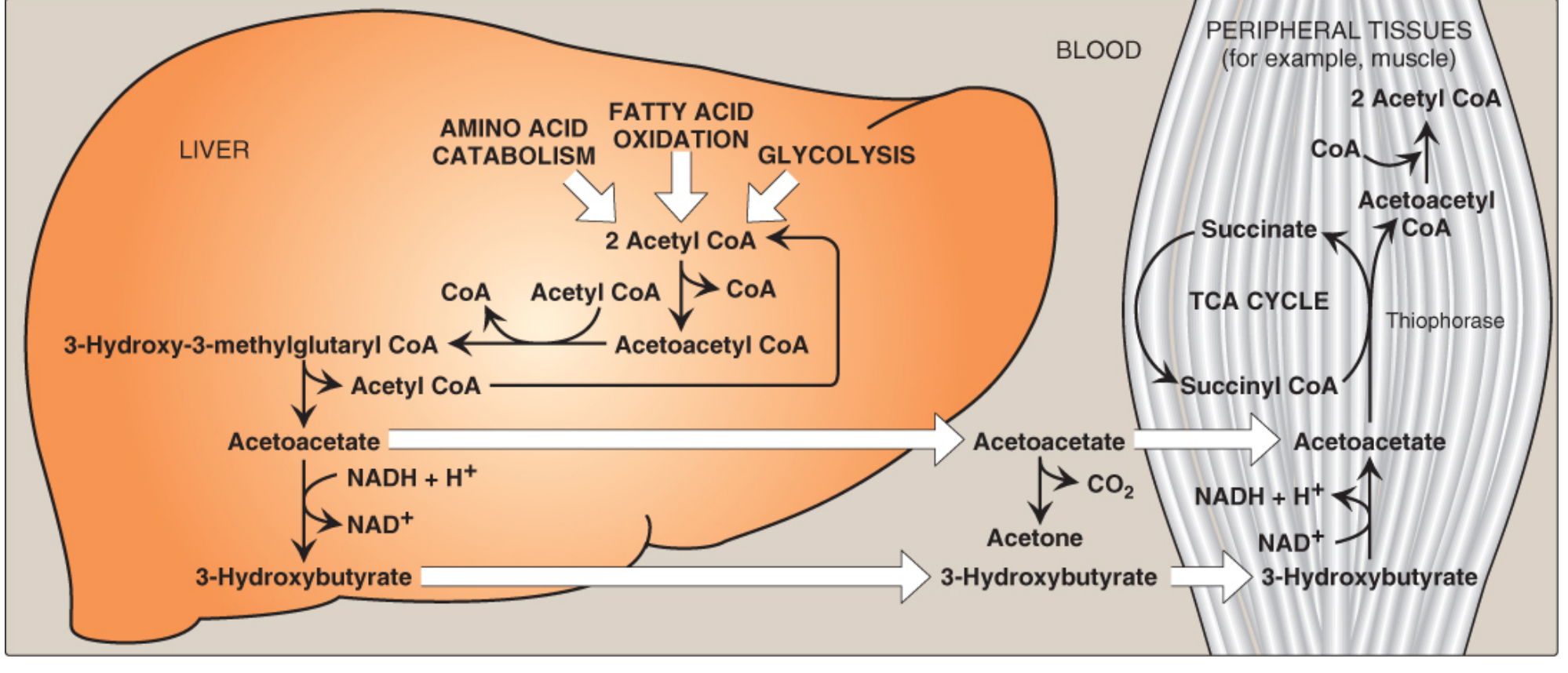
Step-by-step ketolysis:
- 3-Hydroxybutyrate → Acetoacetate: catalyzed by 3-hydroxybutyrate dehydrogenase, generating NADH
- Acetoacetate → Acetoacetyl-CoA: CoA is transferred from succinyl-CoA by thiophorase. This "costs" a succinyl-CoA from the TCA cycle (bypassing the GTP-generating step - one reason the liver cannot afford to do this)
- Acetoacetyl-CoA → 2 Acetyl-CoA: split by thiolase
- 2 Acetyl-CoA → TCA cycle → ATP
Energy yield: 1 mol acetoacetate → 19 ATP; 1 mol 3-hydroxybutyrate → 21.5 ATP
(Biochemistry, Lippincott Illustrated Reviews 8th Ed; Harper's Illustrated Biochemistry 32nd Ed)
Tissue-Specific Reasons
1. Skeletal Muscle
- Muscle preferentially takes up FFAs during fasting, but cannot fully meet energy demands by beta-oxidation alone
- Ketone bodies supplement FFA oxidation when ketone body levels rise in blood
- Importantly, when muscle uses ketone bodies heavily, it reduces its glucose consumption, sparing glucose for the brain and RBCs
- During prolonged starvation, skeletal muscle actually reduces its ketone body uptake (due to low insulin), allowing blood ketone levels to keep rising to eventually supply the brain
(Harper's Illustrated Biochemistry, 32nd Ed, p. 156)
2. Brain
- The brain normally cannot use fatty acids because long-chain fatty acids cannot cross the blood-brain barrier
- Glucose is its usual primary fuel
- However, ketone bodies are small, water-soluble, and freely cross the blood-brain barrier - making them the only fat-derived fuel the brain can use
- After 3-5 days of starvation, the brain starts using ketone bodies significantly; by 7 days, ketone bodies can meet >50% of the brain's energy requirements, reducing glucose demand from ~120 g/day to ~40 g/day
- This is the single most important adaptive advantage: by using ketone bodies, the brain drastically reduces its glucose requirement, so less muscle protein needs to be broken down for gluconeogenesis, sparing body protein and prolonging survival
(Basic Medical Biochemistry 6th Ed, p. 1036; Medical Physiology, p. 1738)
3. Heart Muscle
- The heart is an oxidative powerhouse that runs almost entirely on aerobic metabolism
- It uses ketone bodies very efficiently - heart and skeletal muscle are actually the primary consumers of ketone bodies at moderate ketone levels
- The heart preferentially uses whatever oxidative substrate is most available: fed state → glucose; early fasting → FFAs; prolonged starvation → ketone bodies + FFAs
- Ketone bodies enter directly into the TCA cycle via acetyl-CoA and provide continuous high-efficiency energy for the non-stop cardiac workload
(Harper's Illustrated Biochemistry 32nd Ed; Schwartz's Principles of Surgery 11th Ed)
The Big Picture: Why This Matters for Survival
| Adaptation | Consequence |
|---|---|
| Brain shifts to ketone bodies | Glucose demand falls from ~120 g/day to ~40 g/day |
| Less glucose needed | Hepatic gluconeogenesis decreases |
| Less gluconeogenesis | Less muscle protein is broken down for amino acid substrates |
| Protein is spared | Vital organ function (including the heart) is preserved longer |
| Fat stores are consumed instead | Survival extended from ~10 days (if only protein) to weeks-months |
The shift to ketone body use in these three tissues is therefore a coordinated survival mechanism. It allows the body to "run on fat" while protecting the two most vital protein-rich organs - the brain (which needs a constant fuel source) and the heart (which needs constant oxidative ATP). Starvation deaths typically occur when fat stores are finally depleted and the body must again rely heavily on protein, causing cardiac and diaphragm muscle failure.
(Medical Physiology; Basic Medical Biochemistry 6th Ed)
This is a shared conversation. Sign in to Orris to start your own chat.