Neucleatide Metabolicn
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Finding Sources
Reading File
Reading File
Loading Image
Loading Image
Reading File
Loading Image
I now have comprehensive material. Let me compile the full answer on nucleotide metabolism.
Nucleotide Metabolism
Source: Biochemistry, 8th ed - Lippincott Illustrated Reviews; Basic Medical Biochemistry - A Clinical Approach, 6e
I. Overview
Nucleotides are indispensable to all living cells. Their functions include:
- Building blocks for DNA and RNA synthesis
- Energy currency (ATP, GTP)
- Second messengers (cAMP, cGMP)
- Activated carriers in biosynthesis (UDP-glucose, CDP-choline)
- Coenzyme components (NAD, FAD, CoA)
- Allosteric regulators of metabolic enzymes
Purine and pyrimidine bases can be synthesized de novo (from small molecules) or recycled via salvage pathways. Dietary nucleotides are largely degraded in the GI tract and not significantly utilized.
II. Structure of Nucleotides
A nucleotide = nitrogenous base + pentose sugar + 1-3 phosphate groups.
Purine bases (double ring): Adenine (A), Guanine (G)
Pyrimidine bases (single ring): Cytosine (C), Thymine (T - DNA only), Uracil (U - RNA only)
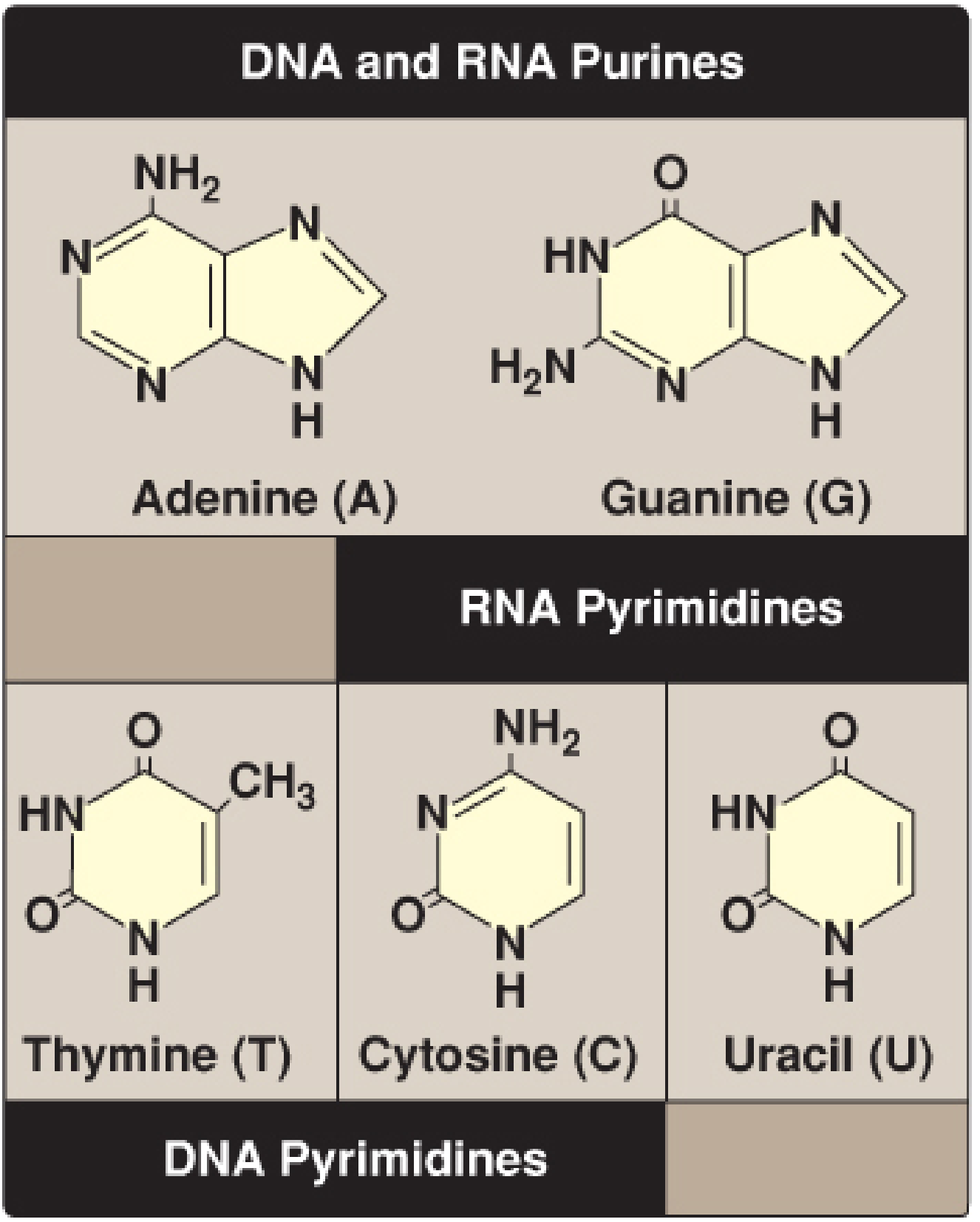
Figure 22.1 - Purines (Adenine, Guanine) and Pyrimidines (Thymine, Cytosine, Uracil). Thymine differs from Uracil by a methyl group at C5. (Lippincott Biochemistry, 8th ed)
III. Purine Synthesis
A. De Novo Purine Synthesis
Purines are synthesized on the ribose phosphate backbone (unlike pyrimidines, which are made as free bases first). The ring is built atom by atom.
Key features:
- Starting material: 5-phosphoribosyl-1-pyrophosphate (PRPP) - formed from ribose 5-phosphate + ATP (by PRPP synthetase)
- Requires 6 ATP per purine synthesized
- First purine product: Inosine monophosphate (IMP)
- AMP and GMP are each derived from IMP in 2-step reactions
Atoms donated to the purine ring:
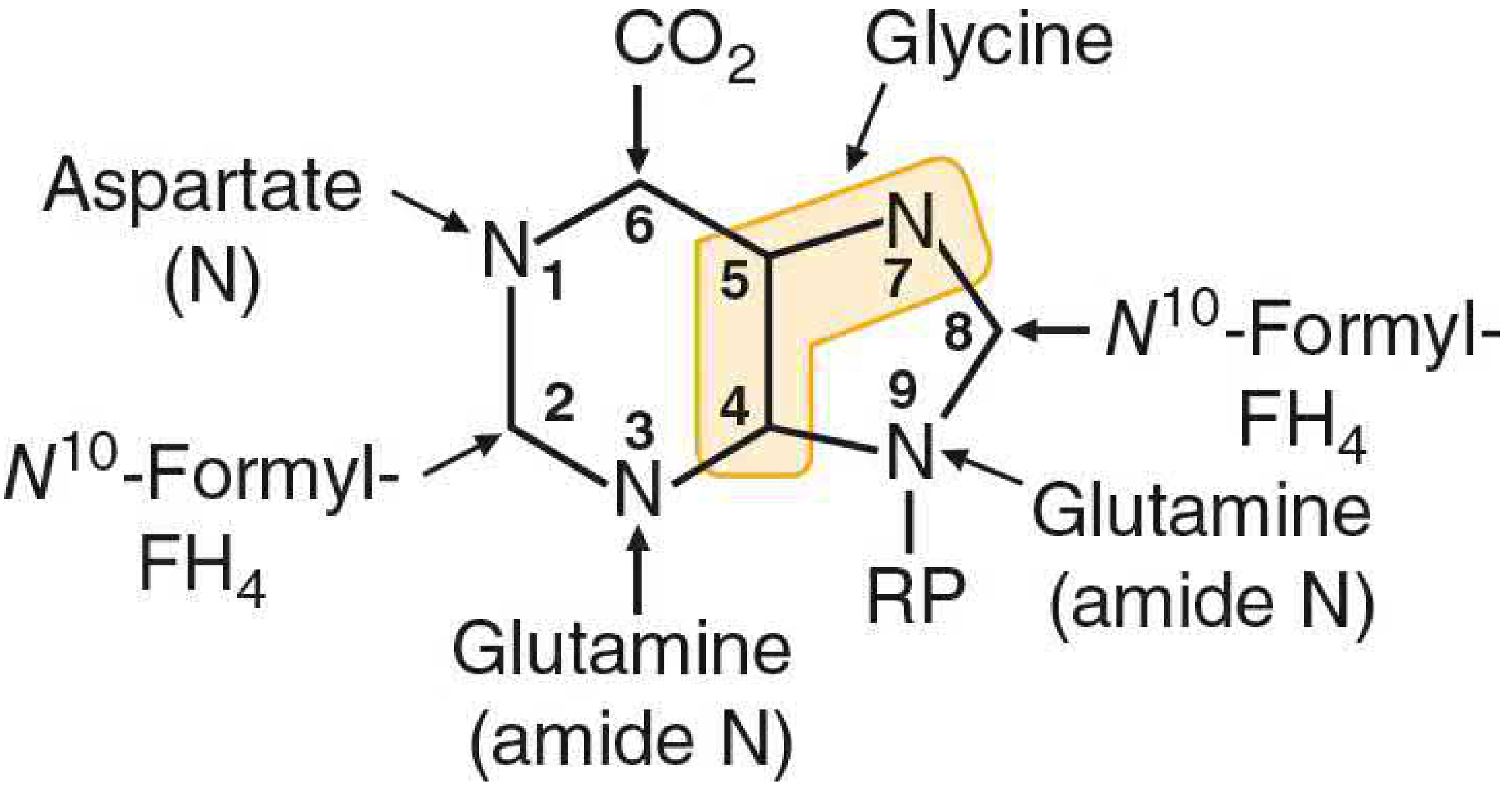
Origin of atoms in the purine ring (Basic Medical Biochemistry, 6e)
| Position | Donor |
|---|---|
| N-1 | Aspartate |
| C-2, C-8 | N10-Formyl-THF (tetrahydrofolate) |
| N-3, N-9 | Glutamine (amide N) |
| C-4, C-5, N-7 | Glycine (entire molecule) |
| C-6 | CO₂ |
Committed step / rate-limiting enzyme: Glutamine phosphoribosylpyrophosphate amidotransferase (GPAT) - converts PRPP + glutamine → 5-phosphoribosylamine
Regulation: AMP and GMP feedback inhibit GPAT (and their respective branches from IMP). PRPP synthetase is feedback inhibited by purine nucleotides.
B. IMP → AMP and IMP → GMP
- IMP → AMP: requires aspartate, GTP (energy); 2 steps
- IMP → GMP: requires NAD⁺ oxidation then glutamine amination, uses ATP (energy)
- Cross-regulation: GTP is used to make AMP; ATP is used to make GMP (balances production)
C. Purine Salvage Pathway
Free purine bases recovered from nucleic acid turnover are recycled to nucleotides. This is far less energy-costly than de novo synthesis.
Key enzymes:
| Enzyme | Reaction |
|---|---|
| HGPRT (hypoxanthine-guanine phosphoribosyltransferase) | Hypoxanthine/Guanine + PRPP → IMP/GMP |
| APRT (adenine phosphoribosyltransferase) | Adenine + PRPP → AMP |
| Adenosine kinase | Adenosine + ATP → AMP + ADP |
Clinical link: Deficiency of HGPRT = Lesch-Nyhan syndrome
IV. Purine Degradation and Clinical Disorders
Purines are degraded to uric acid (the final product in humans, since we lack uricase):
AMP/GMP → Nucleosides → Hypoxanthine/Guanine → Xanthine → Uric acid
The key enzyme is xanthine oxidase (converts hypoxanthine → xanthine → uric acid).
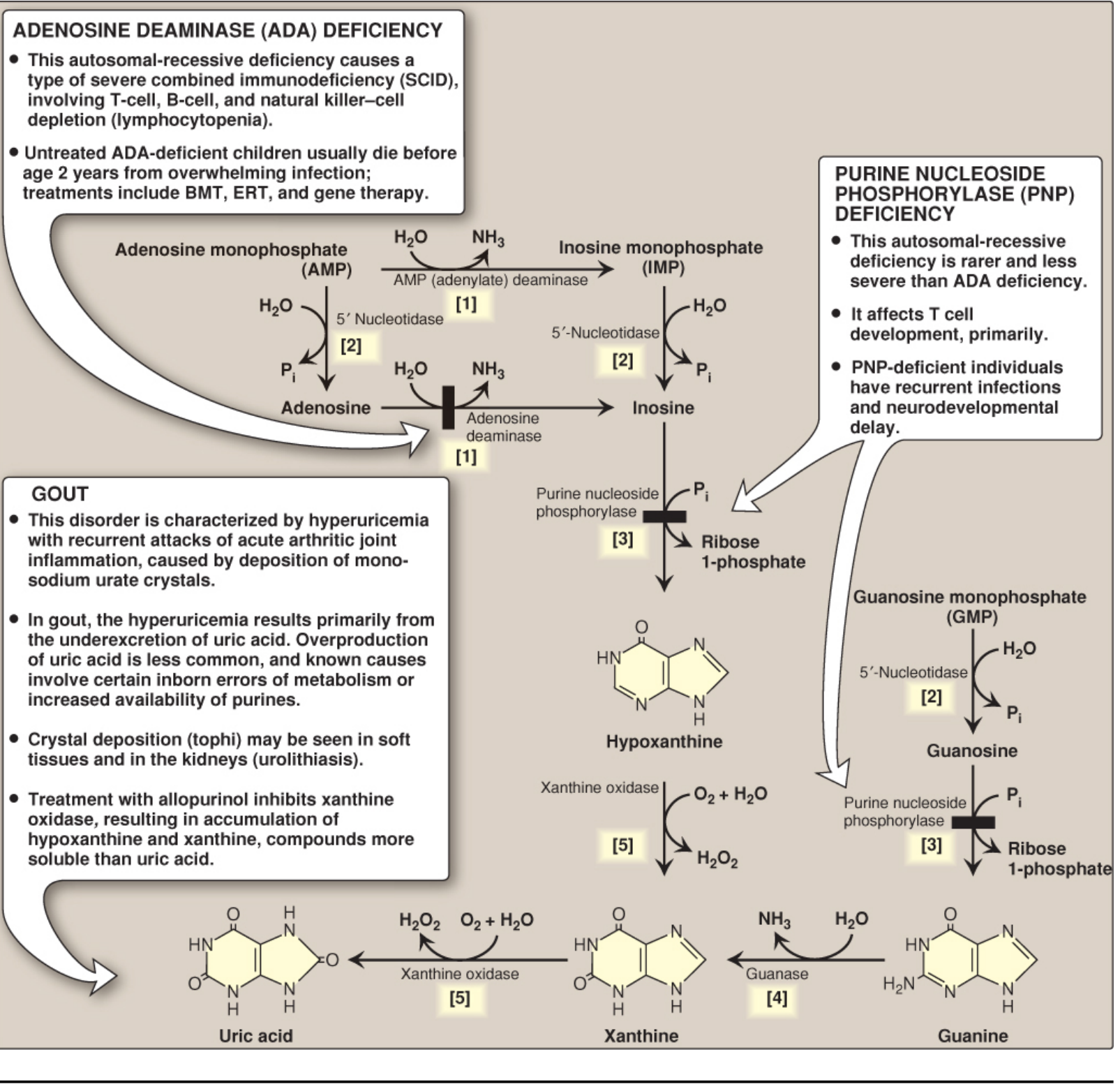
Degradation of purine nucleotides to uric acid with associated genetic disorders (Lippincott Biochemistry, 8th ed)
V. Pyrimidine Synthesis
Pyrimidines are made as a free base first, then attached to PRPP.
De novo synthesis steps:
- Carbamoyl phosphate synthesized by CPS II (cytosolic) using glutamine + CO₂ + 2ATP (note: CPS I is mitochondrial and used in urea cycle)
- Carbamoyl phosphate + aspartate → dihydroorotate → orotate
- Orotate + PRPP → Orotidine 5'-monophosphate (OMP) → UMP
- UMP → UTP → CTP (via CTP synthetase)
- dTMP synthesis: UDP → dUDP → dUMP → dTMP (by thymidylate synthase using N5,N10-methylene-THF)
Committed step: Carbamoyl phosphate synthetase II (CPS II), regulated by PRPP (activator) and UTP (inhibitor)
In mammals, CAD is a trifunctional enzyme with CPS II, aspartate transcarbamoylase, and dihydroorotase activities.
VI. Deoxyribonucleotide Synthesis
Deoxyribonucleotides (needed for DNA) are made from ribonucleoside diphosphates by:
Ribonucleotide reductase (RNR): Reduces ADP, GDP, CDP, UDP → dADP, dGDP, dCDP, dUDP
- Contains two subunits: R1 (α, regulatory) and R2 (β, contains tyrosyl radical)
- Reducing equivalents from thioredoxin → NADPH via thioredoxin reductase (a selenoprotein)
- Allosteric regulation is complex:
- Activity site on R1: dATP inhibits (overall activity); ATP activates
- Specificity site on R1: controls which substrate is used
VII. Clinical Disorders of Nucleotide Metabolism
| Disorder | Enzyme Defect | Mechanism | Features |
|---|---|---|---|
| Gout | Overproduction or underexcretion of uric acid | Hyperuricemia → MSU crystal deposition | Acute/chronic arthritis, tophi, urolithiasis |
| Lesch-Nyhan Syndrome | HGPRT deficiency (X-linked) | Purine salvage failure → ↑ PRPP available → ↑ uric acid + ↑ de novo synthesis | Hyperuricemia, self-mutilation, choreoathetosis, intellectual disability |
| ADA Deficiency | Adenosine deaminase (AR) | Adenosine/dATP accumulate → toxic to lymphocytes | SCID (T, B, NK cell depletion); fatal infection by age 2 if untreated |
| PNP Deficiency | Purine nucleoside phosphorylase (AR) | Purine nucleosides accumulate → T cell death | Primarily T cell immunodeficiency, recurrent infections, neurodevelopmental delay |
| Orotic Aciduria | UMP synthase (UMPS) deficiency | Pyrimidine synthesis blocked; orotic acid accumulates | Megaloblastic anemia, growth retardation; treated with uridine |
| Von Gierke disease | Glucose-6-phosphatase deficiency | ↑ glucose 6-P → ↑ ribose 5-P → ↑ PRPP → ↑ uric acid | Secondary hyperuricemia |
VIII. Pharmacological Targets in Nucleotide Metabolism
| Drug | Target | Use |
|---|---|---|
| Allopurinol | Xanthine oxidase inhibitor | Gout treatment - reduces uric acid production |
| Methotrexate | Dihydrofolate reductase (DHFR) | Cancer, autoimmune disease - blocks THF regeneration, starves cells of purines and dTMP |
| 5-Fluorouracil (5-FU) | Thymidylate synthase | Cancer - blocks dTMP synthesis |
| Hydroxyurea | Ribonucleotide reductase | Cancer, sickle cell disease |
| Mycophenolic acid | IMP dehydrogenase | Immunosuppression - blocks GMP synthesis |
| Probenecid | Renal urate transporter | Gout - increases uric acid excretion |
Quick Summary Table: De Novo Synthesis Comparison
| Feature | Purines | Pyrimidines |
|---|---|---|
| Ring built on | Ribose-5-P (PRPP) | Free base first, then attached to PRPP |
| Committed enzyme | GPAT | CPS II |
| First product | IMP | UMP |
| Atoms donated by | Gly, Asp, Gln, CO₂, N¹⁰-formyl-THF | Asp, Gln, CO₂ |
| Folate required? | Yes (C2, C8) | Yes (dTMP synthesis only) |
| ATP cost | ~6 ATP | ~3 ATP |
Sources: Biochemistry, 8th ed - Lippincott Illustrated Reviews, pp. 821-858 | Basic Medical Biochemistry - A Clinical Approach, 6e, pp. 1406-1415
De Novo Synthesis
Finding Sources
Finding Sources
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Finding Sources
Reading File
Loading Image
Loading Image
Loading Image
Reading File
Reading File
Reading File
I now have comprehensive material for a detailed answer. Let me compile it.
De Novo Synthesis of Nucleotides
Source: Basic Medical Biochemistry - A Clinical Approach, 6e | Biochemistry, 8th ed - Lippincott Illustrated Reviews
PART 1: De Novo Purine Synthesis
Overview
Purines are built atom by atom directly on a ribose-5-phosphate backbone - this is the key distinction from pyrimidines. De novo synthesis occurs primarily in the liver, and to a lesser extent in the brain, neutrophils, and immune cells.
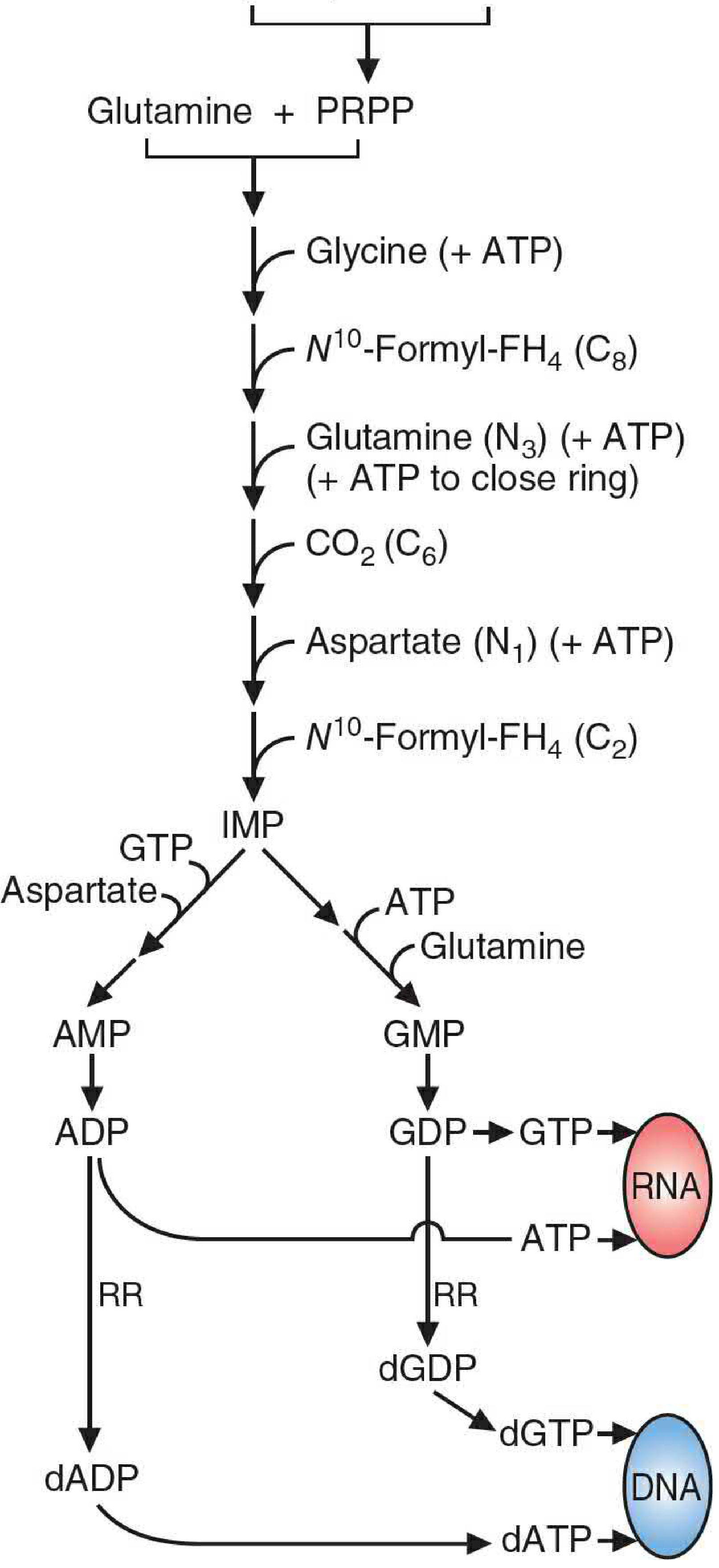
Fig 39.2 - Overview of purine production: Glutamine + PRPP → IMP → AMP/GMP → RNA/DNA (Basic Medical Biochemistry, 6e)
Step-by-Step: The 11 Reactions to IMP
Precursor: PRPP Formation (not the committed step)
Ribose 5-phosphate + ATP → PRPP + AMP
- Enzyme: PRPP synthetase
- Ribose 5-phosphate comes from the pentose phosphate pathway
- PRPP is a shared substrate used in both purine synthesis, pyrimidine synthesis, and salvage pathways
Step 1 - COMMITTED STEP
PRPP + Glutamine → 5-Phosphoribosyl-1-amine + Glutamate + PPi
- Enzyme: Glutamine phosphoribosylpyrophosphate amidotransferase (GPAT)
- This is the first committed, rate-limiting step
- Incorporates N-9 of the purine ring (from glutamine's amide nitrogen)
- The enzyme is a monomer (active) ↔ dimer (inactive) interconversion
- Strongly inhibited by AMP and GMP (end-product feedback inhibition at distinct allosteric sites)
Steps 2-10: Building the Ring Atom by Atom
| Step | What is Added | Atom(s) Donated | Energy |
|---|---|---|---|
| 2 | Glycine added to 5-phosphoribosyl-1-amine | C4, C5, N7 | +ATP |
| 3 | Formyl group from N¹⁰-formyl-THF | C8 | - |
| 4 | Glutamine amide nitrogen | N3 | +ATP |
| 5 | Ring closure (5-membered ring forms) | - | +ATP |
| 6 | CO₂ carboxylation | C6 | - |
| 7 | Aspartate incorporated (then fumarate released) | N1 | +ATP |
| 8 | Formyl group from N¹⁰-formyl-THF | C2 | - |
| 9 | Ring closure → IMP | - | - |
Memory tip for purine ring atoms: "A Genuine Couple Go Alone"
- Aspartate → N1
- Glycine → C4, C5, N7
- CO₂ → C6
- Glutamine → N3, N9
- Alone = N¹⁰-formyl-THF → C2, C8
Total ATP cost to reach IMP: 6 ATP (starting from ribose 5-phosphate)
IMP → AMP (2 steps)
IMP → Adenylosuccinate → AMP
- Adenylosuccinate synthetase: IMP + Aspartate + GTP → Adenylosuccinate + GDP + Pi
- Note: GTP is the energy source (not ATP)
- Adenylosuccinate lyase: Adenylosuccinate → AMP + Fumarate
- Aspartate donates its nitrogen; its carbons leave as fumarate (identical mechanism to urea cycle's argininosuccinate → arginine + fumarate)
IMP → GMP (2 steps)
IMP → XMP → GMP
- IMP dehydrogenase: IMP + NAD⁺ → XMP (xanthosine monophosphate) + NADH
- Oxidation of hypoxanthine base to xanthine
- GMP synthetase: XMP + Glutamine + ATP → GMP + Glutamate + AMP + PPi
- Glutamine donates its amide nitrogen to form the 2-amino group of guanine
Regulation of De Novo Purine Synthesis
AMP ──┐
├──► Inhibit GPAT (committed step)
GMP ──┘
AMP ──► Inhibit adenylosuccinate synthetase (IMP→AMP branch)
GMP ──► Inhibit IMP dehydrogenase (IMP→GMP branch)
ATP ──► needed to make GMP (cross-activation)
GTP ──► needed to make AMP (cross-activation)
This cross-regulation (ATP needed for GMP synthesis; GTP needed for AMP synthesis) balances the relative pool sizes of adenine and guanine nucleotides.
PART 2: De Novo Pyrimidine Synthesis
Overview
Pyrimidines are synthesized differently: the ring is built as a free base first, then attached to PRPP. First product is UMP, which serves as the precursor for all other pyrimidines.
Origin of Pyrimidine Ring Atoms
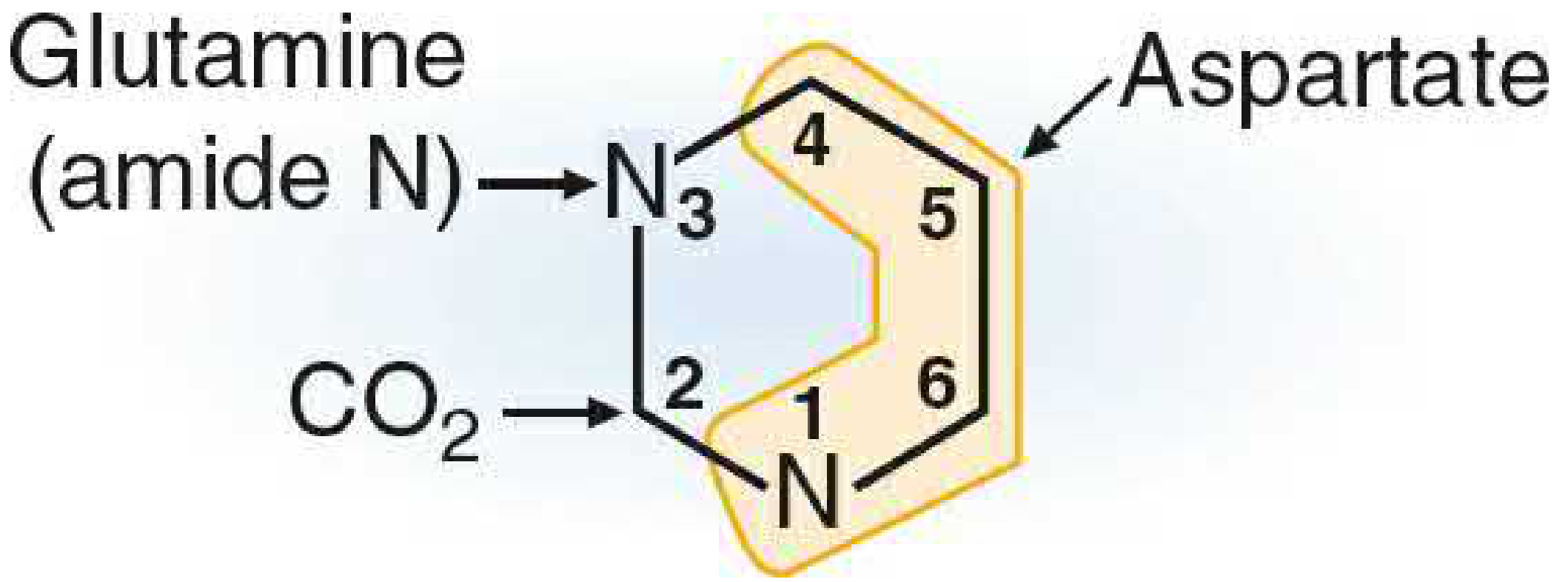
Fig 39.14 - Pyrimidine ring atoms: N1 from Aspartate (amino group), C2 from CO₂, N3 from Glutamine (amide), C4-C6 from Aspartate (Basic Medical Biochemistry, 6e)
| Position | Donor |
|---|---|
| N-1 | Aspartate |
| C-2 | CO₂ |
| N-3 | Glutamine (amide N) |
| C-4, C-5, C-6 | Aspartate |
Only 3 precursor molecules needed: CO₂ + Glutamine + Aspartate (via carbamoyl phosphate)
Step-by-Step: UMP Synthesis (6 steps)
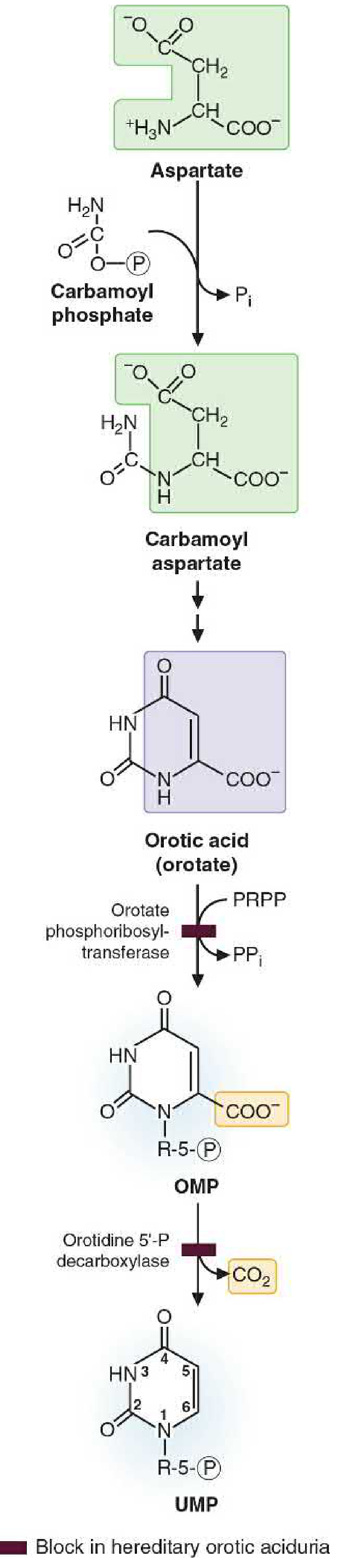
Fig 39.15 - Pyrimidine synthesis: Carbamoyl phosphate + Aspartate → Orotate → OMP → UMP (Basic Medical Biochemistry, 6e)
Step 1 - COMMITTED STEP
Glutamine + CO₂ + 2ATP → Carbamoyl phosphate + Glutamate
- Enzyme: Carbamoyl Phosphate Synthetase II (CPS II) - cytosolic
- This is the committed, rate-limiting step of pyrimidine synthesis
- CPS II is distinct from CPS I (mitochondrial, urea cycle) - different location, different substrate (glutamine vs NH₃), different regulation
- CPS II is part of the CAD trifunctional enzyme
Step 2
Carbamoyl phosphate + Aspartate → Carbamoyl aspartate
- Enzyme: Aspartate transcarbamoylase (part of CAD complex)
Step 3
Carbamoyl aspartate → Dihydroorotate (ring closure)
- Enzyme: Dihydroorotase (part of CAD complex)
CAD = CPS II + Aspartate transcarbamoylase + Dihydroorotase (one polypeptide, 3 activities)
Step 4
Dihydroorotate → Orotate (oxidation)
- Enzyme: Dihydroorotate dehydrogenase - located on the inner mitochondrial membrane (the only mitochondrial step)
- Uses NAD⁺ as electron acceptor
Step 5
Orotate + PRPP → Orotidine 5'-monophosphate (OMP) + PPi
- Enzyme: Orotate phosphoribosyltransferase (part of UMP synthase bifunctional enzyme)
- PRPP attaches the ribose-5-phosphate to the base
Step 6
OMP → UMP + CO₂ (decarboxylation)
- Enzyme: Orotidylic acid decarboxylase (OMP decarboxylase) (part of UMP synthase)
UMP synthase = Orotate phosphoribosyltransferase + OMP decarboxylase (bifunctional enzyme; deficiency causes hereditary orotic aciduria)
UMP → Other Pyrimidines
UMP → UDP → UTP → RNA
↓
CTP synthetase (+Glutamine, +ATP)
↓
CTP → RNA
UDP → dUDP → dUMP → dTMP (via thymidylate synthase + N⁵,N¹⁰-methylene-THF)
→ dTDP → dTTP → DNA
dTMP synthesis is especially important pharmacologically:
- Thymidylate synthase: dUMP + N⁵,N¹⁰-methylene-THF → dTMP + DHF
- DHF is recycled back to THF by dihydrofolate reductase (DHFR)
- Both enzymes are major drug targets (5-FU inhibits thymidylate synthase; methotrexate inhibits DHFR)
Regulation of De Novo Pyrimidine Synthesis
CPS II is the key regulated enzyme:
| Regulator | Effect |
|---|---|
| UTP | Inhibits CPS II (end-product feedback) |
| PRPP | Activates CPS II |
| MAP kinase phosphorylation (S-phase) | Makes enzyme more sensitive to PRPP activation |
| cAMP-dependent protein kinase | Makes enzyme more sensitive to UTP inhibition |
This cell-cycle-linked regulation ensures pyrimidine production is upregulated during DNA synthesis (S-phase).
Clinical note: In ornithine transcarbamoylase (OTC) deficiency (urea cycle disorder), excess mitochondrial carbamoyl phosphate leaks into the cytoplasm, bypasses CPS II regulation, and leads to excessive pyrimidine synthesis - causing orotic aciduria.
PART 3: Comparison Summary
| Feature | Purine (De Novo) | Pyrimidine (De Novo) |
|---|---|---|
| Ring built on | PRPP (ribose backbone) | Free base, then added to PRPP |
| Committed enzyme | GPAT | CPS II |
| Location | Cytosol | Cytosol (except step 4 = mitochondria) |
| First product | IMP (hypoxanthine nucleotide) | UMP |
| Atom donors | Gly, Asp, Gln (×2), CO₂, N¹⁰-formyl-THF (×2) | CO₂, Gln, Asp |
| Folate requirement | Yes (C2 and C8) | Yes (dTMP synthesis only) |
| ATP cost | ~6 ATP per IMP | ~3 ATP per UMP |
| Feedback inhibition | AMP + GMP inhibit GPAT | UTP inhibits CPS II |
| Multifunctional enzymes | Yes (various) | CAD (steps 1-3); UMP synthase (steps 5-6) |
| Key clinical disorder | Lesch-Nyhan, gout | Orotic aciduria |
Key Pharmacological Targets
| Drug | Target | Pathway Blocked |
|---|---|---|
| Methotrexate | Dihydrofolate reductase (DHFR) | Blocks THF regeneration → no purine C2/C8 or dTMP |
| 5-Fluorouracil (5-FU) | Thymidylate synthase | Blocks dTMP synthesis |
| Hydroxyurea | Ribonucleotide reductase | Blocks deoxyribonucleotide formation |
| Azaserine / DON | Glutamine-utilizing enzymes | Inhibits multiple steps (GPAT, GMP synthetase) |
| Mycophenolic acid | IMP dehydrogenase | Blocks GMP synthesis |
| Leflunomide | Dihydroorotate dehydrogenase | Blocks pyrimidine synthesis (Step 4) |
Sources: Basic Medical Biochemistry - A Clinical Approach, 6e, pp. 1408-1432 | Biochemistry, 8th ed - Lippincott Illustrated Reviews, pp. 821-858
This is a shared conversation. Sign in to Orris to start your own chat.