I now have comprehensive, well-sourced content with excellent diagrams. Let me compile the full, detailed answer.
Physiology of Hearing
1. The Nature of Sound
Sound is a form of mechanical energy consisting of alternating compressions and rarefactions propagating through an elastic medium. In air at 20°C at sea level, sound travels at approximately 344 m/s (some sources cite 340 m/s). Sound travels faster through liquids and solids. When sound energy must cross from air to a fluid medium, nearly 99.9% of the acoustic energy is reflected due to the difference in acoustic impedance between the two media - this is the fundamental engineering problem the ear must solve.
2. Overview: Three Functional Divisions
The mechanism of hearing is divided into three sequential stages:
| Stage | Structures | Function |
|---|
| Conductive | Pinna, ear canal, tympanic membrane, ossicles | Capture and amplify sound; impedance matching |
| Sensory (Transduction) | Cochlea, organ of Corti, hair cells | Convert mechanical energy to electrical signals |
| Neural | CN VIII, central auditory pathways | Transmit and process signals to cortex |
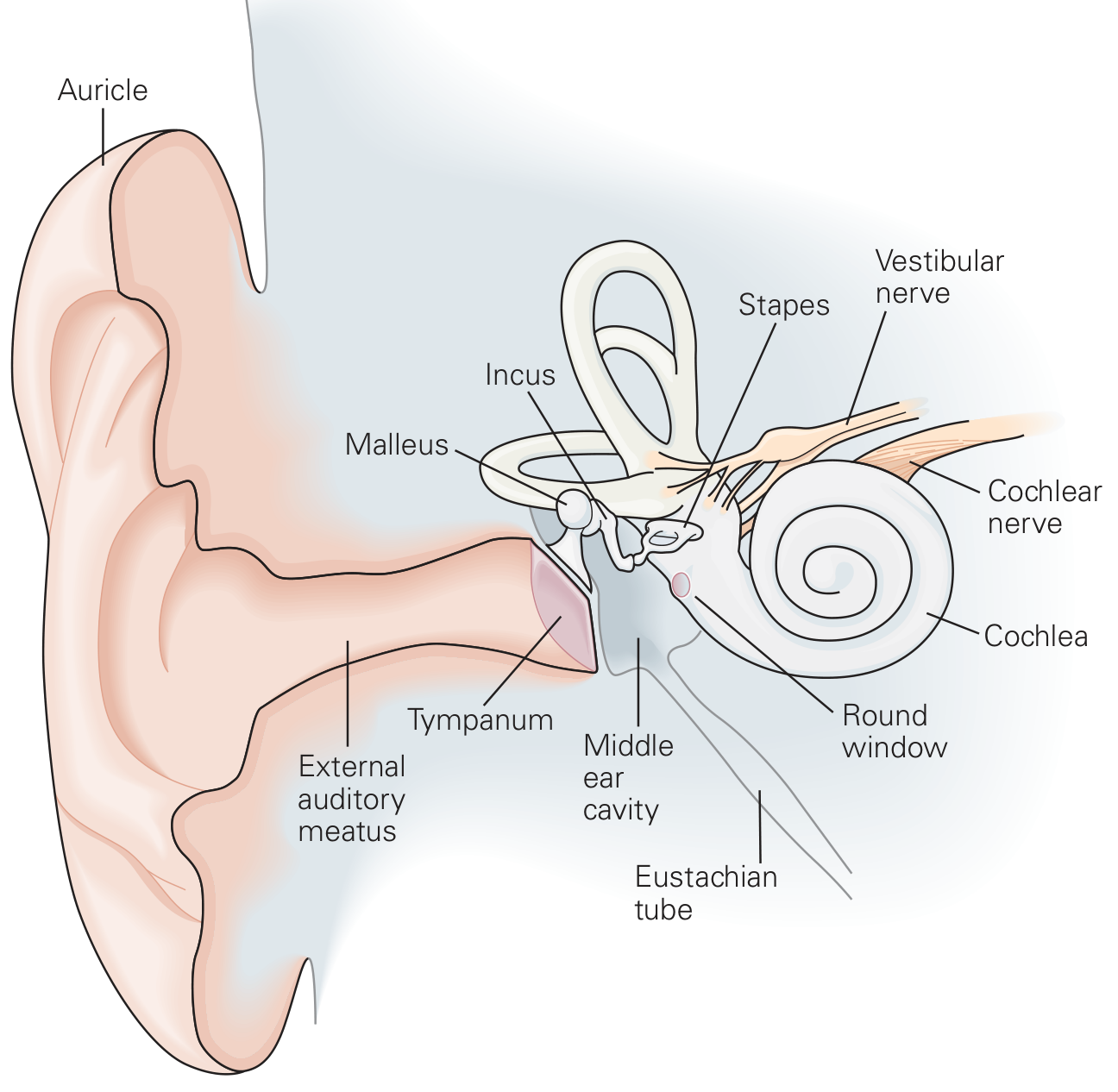
3. Anatomy of the Ear
Fig. 1 - Structure of the human ear (Kandel, Principles of Neural Science)
External ear:
- The auricle (pinna) is a cartilage-supported skin fold that captures sound and directs it into the external auditory meatus. Its corrugated surface is important for vertical sound localization - each person's auricle has a unique topography whose effect on sound reflection is learned early in life.
- The external auditory canal ends at the tympanic membrane (diameter ~9 mm, thickness ~50 µm).
- Natural resonance of the external auditory canal: ~1000 Hz
Middle ear:
- An air-filled pouch connected to the nasopharynx via the Eustachian tube (equalizes pressure).
- Contains three ossicles: malleus (hammer), incus (anvil), stapes (stirrup) - linked in a chain.
- The handle of the malleus is attached to the tympanic membrane; the footplate of the stapes sits in the oval window of the cochlea.
- Natural resonance of the middle ear: ~800 Hz; ossicular chain most efficient at 500-2000 Hz; tympanic membrane most efficient at 800-1600 Hz.
Inner ear (cochlea):
- A coiled, snail-shaped bony structure (~9 mm across in humans), embedded in the temporal bone.
- Three fluid-filled chambers:
- Scala vestibuli (top) - perilymph; begins at the oval window
- Scala media (middle) - endolymph (high K+, low Na+, similar to intracellular fluid)
- Scala tympani (bottom) - perilymph; ends at the round window
- Scala vestibuli is separated from scala media by Reissner's membrane.
- Scala media is separated from scala tympani by the basilar membrane.
4. Impedance Matching: The Transformer Action of the Middle Ear
The key problem: sound energy traveling from air to cochlear fluid would lose 99.9% at the interface. The middle ear compensates by acting as an impedance-matching transformer, converting sound of large amplitude/low force into sound of small amplitude/high force. This is accomplished by three mechanisms:
(a) Hydraulic (Area) Ratio of the Tympanic Membrane
The tympanic membrane (~90 mm² total, ~55 mm² functional) is far larger than the stapes footplate (~3.2 mm²).
- Effective areal ratio = ~14:1 (using two-thirds of the TM as the functional area)
- Per Wever and Lawrence: 55 mm² functional TM / 3.2 mm² footplate = 17:1
(b) Lever Action of the Ossicular Chain
The handle of the malleus is 1.3 times longer than the long process of the incus, providing a mechanical advantage of 1.3:1.
(c) Curved Membrane Effect
The tympanic membrane moves more at its periphery than at its center (where the malleus is attached), providing additional leverage.
Total transformer ratio:
- Basic: 14 × 1.3 = ~18:1
- Per Wever and Lawrence: 17 × 1.3 = ~22:1
- Harrison's 22nd ed. states the eardrum and ossicles boost sound energy ~200-fold (accounting for all mechanical effects combined).
Phase Differential Between Oval and Round Windows
The ossicular chain creates a preferential pathway to the oval window - so when the oval window receives a compression wave, the round window is in the rarefaction phase. This out-of-phase relationship is essential: if both windows received simultaneous pressure waves, they would cancel each other out with no net movement of perilymph and no hearing. This acoustic separation requires an intact tympanic membrane and air in the middle ear space around the round window. The phase differential contributes approximately 4 dB when the tympanic membrane is intact.
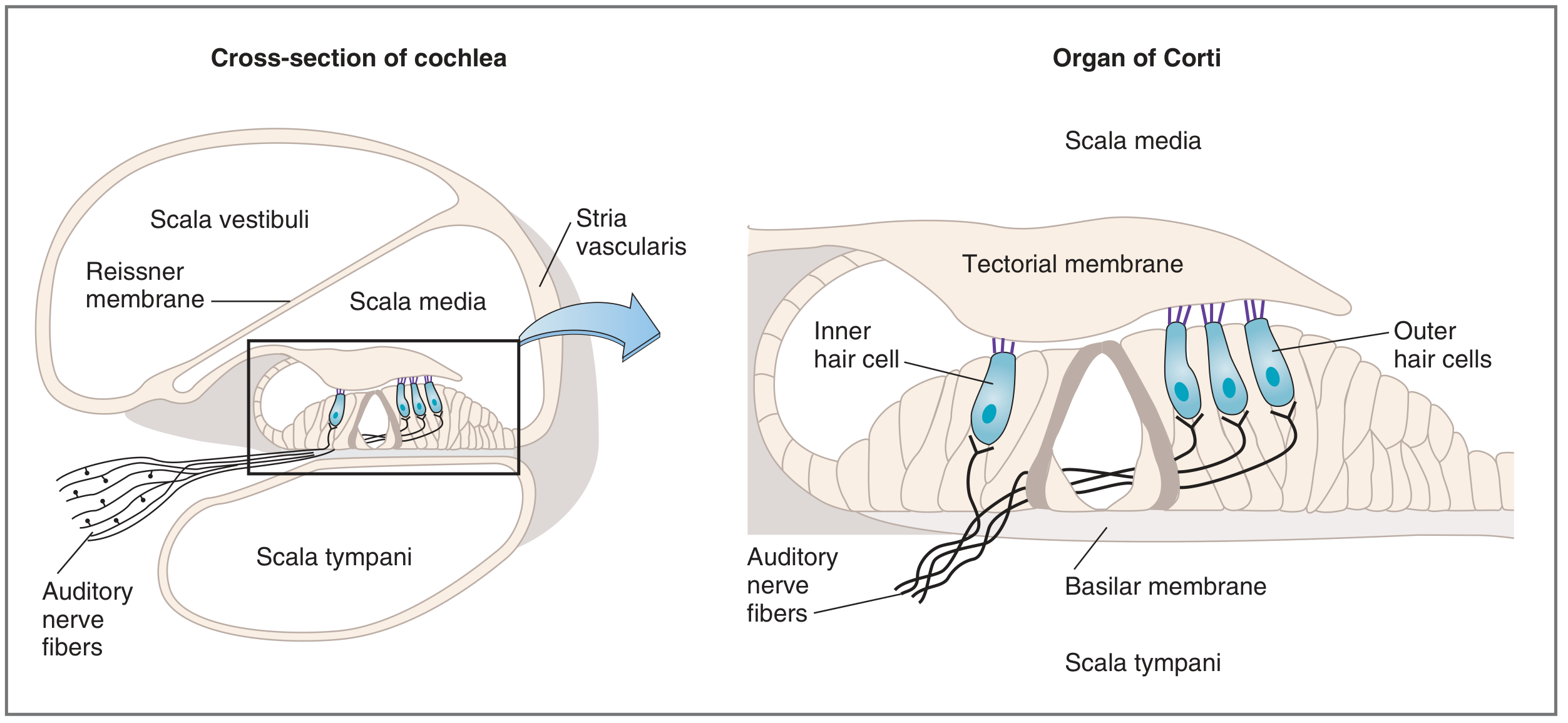
5. Cochlear Anatomy and the Organ of Corti
Fig. 2 - Cross-section of the cochlea and the organ of Corti (Costanzo Physiology, 7th ed.)
The organ of Corti lies on the basilar membrane, bathed in endolymph of the scala media. It contains two types of mechanoreceptor hair cells:
| Feature | Inner Hair Cells | Outer Hair Cells |
|---|
| Arrangement | Single row | 3 parallel rows |
| Number | ~3,500 | ~20,000 |
| Innervation | Primarily afferent (95% of CN VIII fibers) | Primarily efferent |
| Function | Principal sensory transducers | Cochlear amplifier (electromotility) |
The cilia (stereocilia) of hair cells are embedded in the tectorial membrane above, while their cell bodies sit on the basilar membrane below. The tectorial membrane is stiffer/less elastic than the basilar membrane.
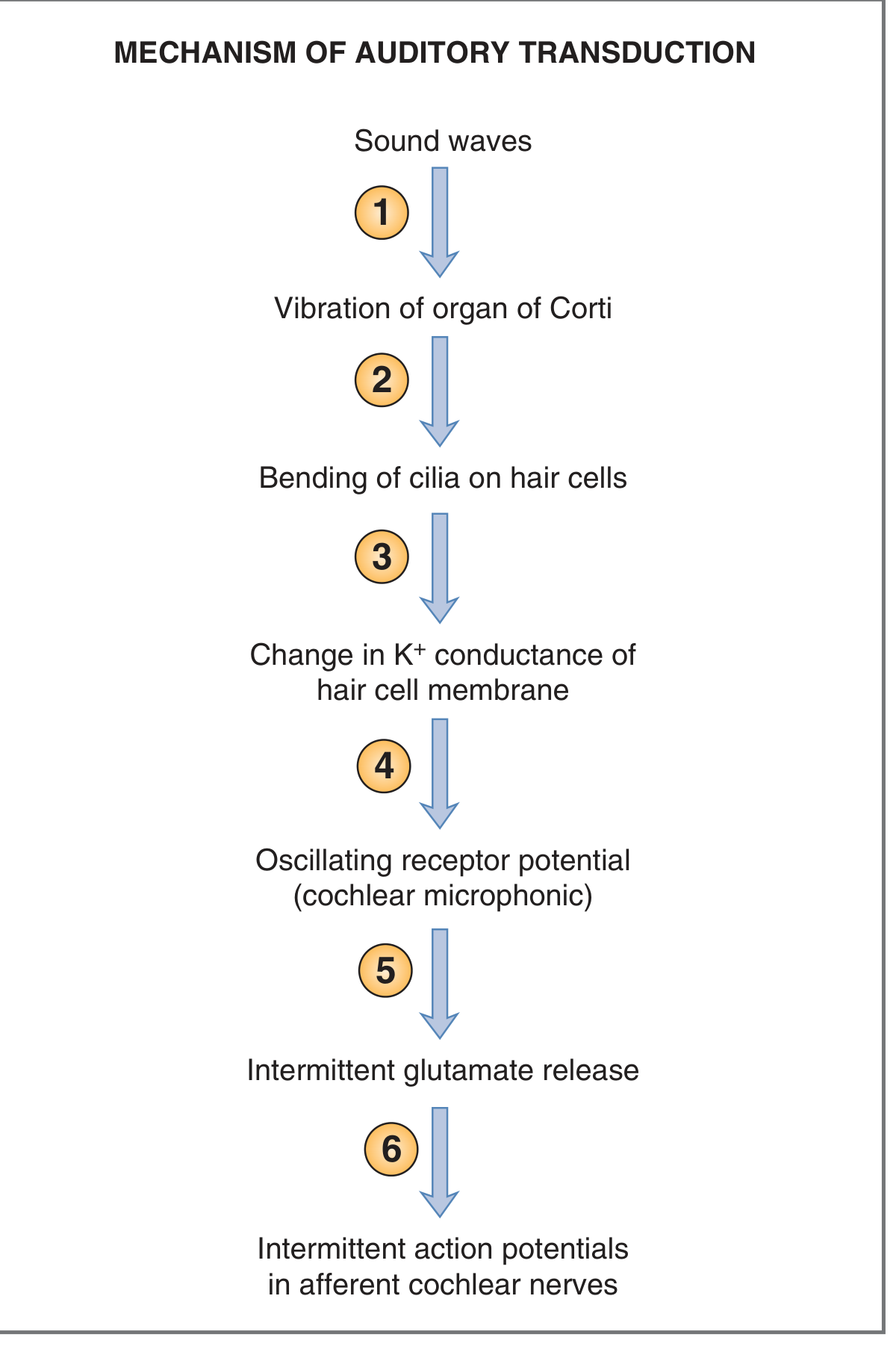
6. Mechanoelectrical Transduction: Step-by-Step
Fig. 3 - Steps in auditory transduction (Costanzo Physiology, 7th ed.)
- Sound waves arrive at the tympanic membrane and are transmitted via the ossicular chain to the stapes footplate at the oval window.
- Stapes movements create pressure waves in perilymph - the scala vestibuli pressure rises relative to scala tympani, causing the basilar membrane to bow downward.
- The basilar membrane vibrates - because it is more elastic than the tectorial membrane, vibration creates a shearing force between the basilar membrane (moving) and the tectorial membrane (more stationary), bending the stereocilia.
- Stereocilia deflection - bending toward the tallest stereocilium stretches tip links (fine filamentous connections between adjacent stereocilia), mechanically pulling open K+ channels (MET channels). K+ floods into the hair cell from endolymph (which has unusually high K+ concentration, similar to intracellular fluid). This causes depolarization - a receptor potential called the cochlear microphonic (CM).
- Depolarization opens voltage-gated Ca²+ channels in the presynaptic terminal of the hair cell.
- Ca²+ entry triggers vesicular release of glutamate at the ribbon synapse, generating action potentials in afferent cochlear nerve fibers.
Note on K+ paradox: Because hair cell cilia are bathed in endolymph (high K+), K+ flows into the cell when channels open - the opposite of what happens in most neurons. Bending toward the tallest stereocilium = depolarization; bending away = hyperpolarization.
Outer hair cell role (cochlear amplifier): The motility of outer hair cells (driven by the protein prestin in their lateral wall) physically amplifies basilar membrane motion, providing the cochlea with its exquisite sensitivity (~10 nm motion) and sharp frequency tuning. This active amplification mechanism explains why the cochlea is so much more sensitive than a passive system.
7. Tonotopic Organization and the Travelling Wave (von Bekesy)
According to Georg von Bekesy's travelling wave theory (Nobel Prize 1961), a sound wave entering the cochlea does not displace the entire basilar membrane equally. Instead, it creates a travelling wave that propagates from the base toward the apex, reaching its point of maximum amplitude at a location determined by the sound's frequency:
- High frequencies (e.g., 20,000 Hz) → maximum displacement near the base (near the stapes)
- Low frequencies (e.g., 20-200 Hz) → maximum displacement near the apex
- Speech-range frequencies (~500-4000 Hz) → represented at intermediate positions
This is the basis of tonotopic (place) coding - the cochlea acts as a mechanical frequency analyzer. The basilar membrane is narrow and stiff at the base, wide and floppy at the apex, which is what creates this frequency selectivity.
8. Electrical Potentials of the Cochlea
Four types of electrical potentials are recorded from the cochlea and CN VIII:
(1) Endocochlear Potential (EP)
- A DC potential of +80 mV recorded from the scala media.
- Generated by the stria vascularis via the Na+/K+-ATPase pump.
- Present at rest (no sound stimulus required).
- Acts as a "battery" - provides the electrochemical driving force for K+ influx into hair cells during transduction.
- The resting potential inside the hair cell is approximately -40 to -70 mV, so the combined driving force across the stereocilia tip is +80 mV + ~60 mV = ~120-140 mV (Davis' battery model).
(2) Cochlear Microphonic (CM)
- An AC (alternating current) potential generated by hair cells.
- Mirrors the waveform of the stimulating sound (like a microphone, hence the name).
- Results from oscillating K+ conductance changes as the basilar membrane vibrates.
- A receptor potential - graded (not all-or-none), no latency, not propagated, no refractory period.
(3) Summating Potential (SP)
- A DC potential that follows the "envelope" of the stimulating sound.
- Generated by hair cells; can be positive or negative.
- Also a receptor potential (shares properties with CM above).
- Clinically used in the electrocochleography (ECoG) diagnosis of Ménière's disease (elevated SP/AP ratio is a key diagnostic marker).
(4) Compound Action Potential (AP / CAP)
- An all-or-none response of the auditory nerve fibers.
- Reflects the summed firing of many CN VIII fibers.
- Has latency, is propagated, follows all properties of a true nerve action potential.
9. Central Auditory Pathways
Hair cells synapse on bipolar neurons of the spiral ganglion (located in the modiolus of the cochlea). Their central axons form the cochlear nerve (CN VIII). The auditory pathway then ascends:
Spiral ganglion (CN VIII)
↓
Cochlear nuclei (ventral + dorsal) — first synapse, in medulla
↓ (both crossed and uncrossed fibers)
Superior olivary nucleus (pons) — key for sound localization/binaural processing
↓
Lateral lemniscus
↓
Inferior colliculus (midbrain) — reflex responses to sound
↓
Medial geniculate body (thalamus)
↓
Auditory cortex (Heschl's gyri, Brodmann's area 41)
— located at the middle of the superior temporal gyrus
Key features of the central pathway:
- Tonotopic organization is preserved at every level from cochlea to cortex.
- Both crossed and uncrossed fibers exist at multiple levels, meaning each cortex receives input from both ears.
- At low frequencies, auditory nerve fibers fire synchronously with the sound wave (phase locking). At higher frequencies, neurons alternate their firing across phases.
- Sound intensity is encoded by: firing rate of individual neurons, total number of active neurons, and which specific neurons are activated.
Cortical lateralization:
- The left auditory cortex is specialized for speech recognition and production.
- The right hemisphere processes emotional and tonal aspects of speech.
- Left hemisphere dominance for speech: found in 95-98% of right-handed persons and 70-80% of left-handed persons.
- A "right ear advantage" exists for dichotic listening tasks involving consonants and words - because right-ear signals cross to the dominant left hemisphere.
10. Summary of Frequency Transmission Efficiency
| Structure | Best Frequency Range |
|---|
| External auditory canal | ~1000 Hz (natural resonance) |
| Tympanic membrane | 800-1600 Hz |
| Middle ear (overall) | 800 Hz (resonance) |
| Ossicular chain | 500-2000 Hz |
| Overall best range | 500-3000 Hz |
This 500-3000 Hz range corresponds precisely to the frequencies most important for human speech, reflecting evolutionary optimization of the auditory apparatus.
Sources:
- Harrison's Principles of Internal Medicine, 22nd ed. (2025), Chapter 36 - Physiology of Hearing
- Costanzo Physiology, 7th ed. - Auditory Transduction; Cochlea and Organ of Corti
- Kandel, Principles of Neural Science, 6th ed. - Chapter 26: Hearing
- The passage provided (Scott-Brown/ENT textbook, Chapter 2)