Hii
Hey! How can I help you today?
Translation in eukayotes
Reading File
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
I now have comprehensive content. Let me compile the full answer.
Translation in Eukaryotes
Translation is the process of synthesizing a protein from the nucleotide sequence of an mRNA molecule. In eukaryotes, this occurs in the cytoplasm on 80S ribosomes (composed of a large 60S and a small 40S subunit). Unlike prokaryotes, eukaryotic transcription occurs in the nucleus while translation occurs in the cytoplasm, so the two processes are never simultaneous. Each eukaryotic mRNA codes for only one protein (it is monocistronic).
Key Components Required
| Component | Role |
|---|---|
| 80S ribosome (40S + 60S subunits) | Machinery of translation |
| mRNA (with 5' cap and poly-A tail) | Template |
| Aminoacyl-tRNA (aa-tRNA) | Brings amino acids |
| Eukaryotic Initiation Factors (eIFs) | Assemble the initiation complex |
| Eukaryotic Elongation Factors (eEFs) | Facilitate peptide chain elongation |
| Eukaryotic Release Factors (eRFs) | Terminate translation |
| GTP and ATP | Energy sources |
Stage 1: Initiation
This is the most complex and regulated stage. It involves assembly of the ribosome on the mRNA before peptide bond formation.
Steps:
1. Ribosome Dissociation
- The 80S ribosome must first dissociate into its 40S and 60S subunits.
- Initiation factors eIF-1, eIF-1A, and eIF-3 bind the newly free 40S subunit and prevent premature reassociation with 60S.
2. Formation of the 43S Preinitiation Complex
- eIF-2-GTP binds the special initiator Met-tRNA (Met-tRNA_i) to form a ternary complex.
- This ternary complex joins the 40S subunit (along with eIF-1, eIF-1A, eIF-3, and eIF-5) to form the 43S preinitiation complex.
- Note: In eukaryotes, the initiator tRNA carries unmodified methionine (Met) - not formylmethionine (fMet) as in prokaryotes.
3. mRNA Activation and 48S Initiation Complex
- The mRNA is recognized through its 5' cap structure (m7G cap).
- eIF-4E (cap-binding protein) binds the 5' cap.
- eIF-4G (scaffold protein) and eIF-4A (an ATP-dependent RNA helicase) join eIF-4E to form the eIF-4F complex.
- Poly-A binding proteins (PABPs) interact with the poly-A tail and bind to eIF-4G, causing circularization of the mRNA - this likely prevents use of incompletely processed mRNAs.
- The 43S complex binds near the 5' cap and scans 5'→3' along the mRNA (this scanning requires ATP hydrolysis by eIF-4A) until it finds the first AUG codon in a favorable Kozak sequence context.
- This produces the 48S initiation complex.
Cap-independent initiation: Some mRNAs (e.g., viral mRNAs) use an Internal Ribosome Entry Site (IRES) that allows the 40S subunit to bind directly near the start codon without scanning.
4. 80S Initiation Complex Formation
- eIF-5 triggers GTP hydrolysis on eIF-2, releasing eIF-2-GDP and several other eIFs.
- The large 60S subunit then joins with the help of eIF-5B-GTP, forming the complete 80S initiation complex.
- The initiator Met-tRNA now occupies the P site of the ribosome, and the A site is empty and ready.
Stage 2: Elongation
The ribosome moves along the mRNA in the 5'→3' direction, adding one amino acid per codon. Three ribosomal sites are involved:
- A (Aminoacyl) site - accepts incoming aa-tRNA
- P (Peptidyl) site - holds the growing polypeptide chain
- E (Exit) site - releases the discharged tRNA
Steps:
1. Aminoacyl-tRNA Binding (A site occupation)
- eEF-1A-GTP (equivalent to prokaryotic EF-Tu) delivers the correct aminoacyl-tRNA to the A site.
- Codon-anticodon recognition triggers GTP hydrolysis; eEF-1A-GDP is released.
- eEF-1B (equivalent to EF-Ts) exchanges GDP for GTP to regenerate active eEF-1A.
2. Peptide Bond Formation (Transpeptidation)
- The peptidyl transferase activity of the 23S rRNA in the 60S subunit (a ribozyme activity) catalyzes formation of a peptide bond between the amino acid in the A site and the growing chain in the P site.
- The chain is transferred from P-site tRNA to the A-site tRNA; the P-site tRNA is now uncharged.
3. Translocation
- eEF-2-GTP (equivalent to prokaryotic EF-G) catalyzes movement of the ribosome one codon (3 nucleotides) in the 3' direction.
- The peptidyl-tRNA moves from A→P site, the empty tRNA moves from P→E site (and exits), and the A site is now empty for the next codon.
- GTP is hydrolyzed in this step.
This cycle repeats for each codon until a stop codon is reached.
Stage 3: Termination
1. Stop Codon Recognition
- When a stop codon (UAA, UAG, or UGA) enters the A site, no tRNA recognizes it.
- Instead, eRF-1 (a eukaryotic release factor that mimics a tRNA structure) recognizes all three stop codons.
- eRF-3-GTP works with eRF-1 to stimulate release.
2. Polypeptide Release
- eRF-1 triggers the peptidyl transferase to add water instead of an amino acid, hydrolyzing the bond between the polypeptide and the tRNA.
- The completed polypeptide is released from the ribosome.
3. Ribosome Recycling
- The 80S ribosome dissociates into 40S and 60S subunits, which can be reused.
Polysomes (Polyribosomes)
Multiple ribosomes can translate a single mRNA molecule simultaneously, forming a structure called a polysome. This greatly increases the efficiency of protein synthesis.
Co- and Post-Translational Modifications
After translation, proteins often undergo modifications:
- Trimming - removal of signal peptides or precursor sequences (e.g., insulin, procollagen)
- Phosphorylation - on Ser, Thr, or Tyr residues by protein kinases
- Glycosylation - N-linked (in RER) or O-linked (in Golgi) sugars added
- Acetylation, Hydroxylation, Ubiquitination, and more
Comparison: Eukaryotic vs Prokaryotic Translation
| Feature | Eukaryotes | Prokaryotes |
|---|---|---|
| Ribosome | 80S (40S + 60S) | 70S (30S + 50S) |
| mRNA type | Monocistronic | Polycistronic |
| Start codon recognition | 5' cap scanning | Shine-Dalgarno sequence |
| Initiator tRNA | Met-tRNA_i | fMet-tRNA_i |
| Initiation factors | Many eIFs (>10) | 3 IFs |
| Coupled transcription/translation | No (nuclear membrane separates) | Yes |
| Location | Cytoplasm (also mitochondria) | Cytoplasm |
| Energy for initiation | GTP + ATP | GTP only |
Sources: Biochemistry (Lippincott Illustrated Reviews, 8th ed), pp. 1254-1265 | Harper's Illustrated Biochemistry, 32nd ed, pp. 421+
Wooble hypothesis
Finding Sources
Reading File
Loading Image
Loading Image
Here is a complete explanation of the Wobble Hypothesis:
The Wobble Hypothesis
Proposed by: Francis Crick, 1966
Background - Why Wobble is Needed
The genetic code has 64 codons (61 sense codons + 3 stop codons). If one unique tRNA were required for each sense codon, 61 different tRNAs would be needed. However, the actual number of tRNAs in a cell is far fewer (around 40-45 in humans). The wobble hypothesis explains how a single tRNA can recognize multiple synonymous codons.
Core Concept
Codon-anticodon pairing is antiparallel - the mRNA codon is read 5'→3', and the tRNA anticodon pairs in the opposite 3'→5' direction.
The hypothesis states:
-
Positions 1 and 2 of the codon (5' end of codon) follow strict, traditional Watson-Crick base pairing with positions 3 and 2 of the anticodon:
- A pairs with U
- G pairs with C
- U pairs with A
- C pairs with G
-
Position 3 of the codon (3' end = the "wobble position") can form non-traditional, flexible pairings with the 5' base of the anticodon (position 1 of anticodon). This flexibility is called wobble.
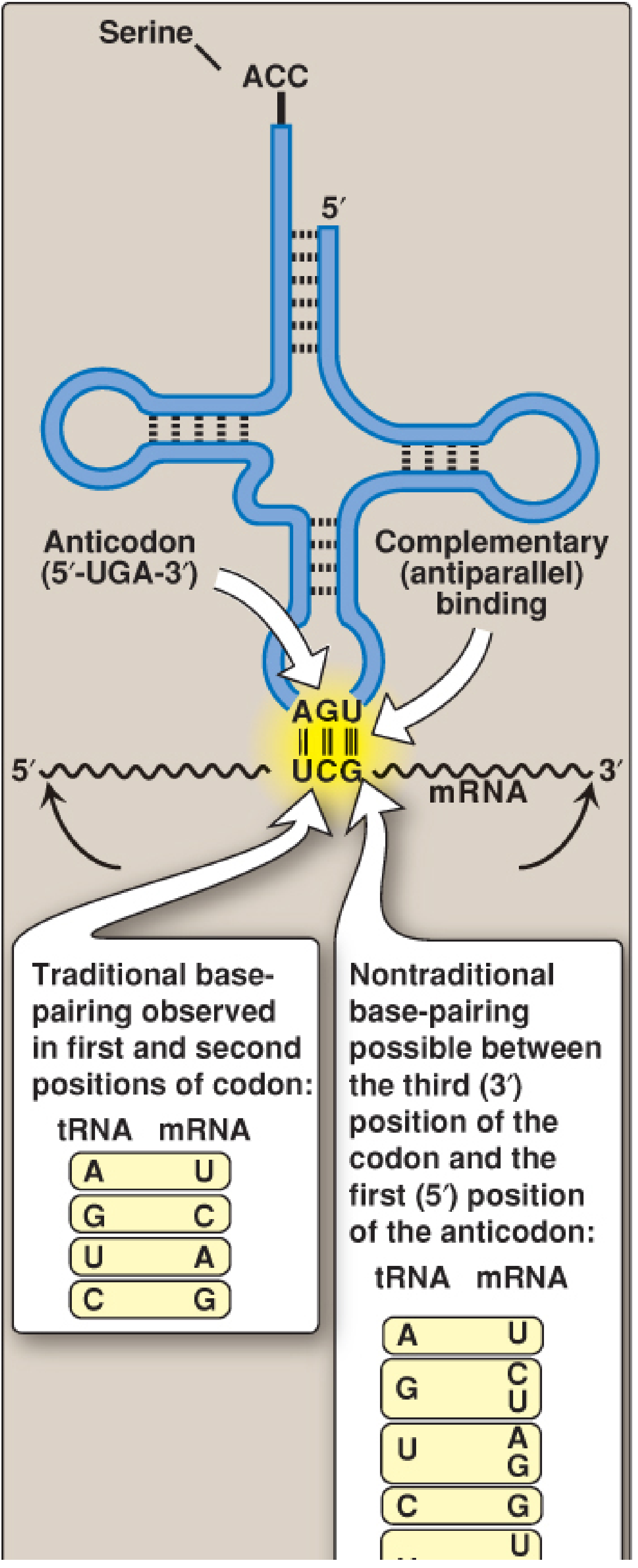
Figure 32.9 from Biochemistry, Lippincott Illustrated Reviews, 8th ed - showing traditional pairing at positions 1 & 2 of the codon (left box) vs. nontraditional wobble pairing at position 3 (right box)
Wobble Base Pairings
The 5' base of the anticodon (wobble position) can pair with the following bases at the 3' position of the codon:
| 5' Base of Anticodon (tRNA) | 3' Base of Codon (mRNA) it can pair with |
|---|---|
| A | U only |
| G | C or U |
| U | A or G |
| C | G only |
| I (Inosine) | U, C, or A |
Inosine (I) is a modified base formed by deamination of adenine (its base is hypoxanthine). It is the most promiscuous wobble base - a tRNA with inosine at the 5' anticodon position can read three different codons.
Significance of Wobble
- Fewer tRNAs needed - 61 tRNAs are NOT required for 61 codons; ~40-45 tRNAs suffice.
- Explains degeneracy of the genetic code - Multiple codons can specify the same amino acid, and wobble explains how one tRNA handles this.
- Synonymous codons often differ only at the 3rd position - This is the "third base degeneracy" or "wobble position" seen throughout the genetic code (e.g., Gly: GGU, GGC, GGA, GGG - one tRNA with anticodon CCC can read GGG, and one with anticodon ICC can read GGU, GGC, and GGA).
- Inosine-containing tRNAs are especially efficient - A single tRNA with inosine at the wobble position can decode three codons.
Simple Summary
mRNA Codon: 5' - N1 - N2 - N3(wobble) - 3'
| | |
tRNA Anticodon: 3' - N1' - N2' - N3'(wobble) - 5'
Positions 1 & 2 → Strict Watson-Crick pairing
Position 3 (3' of codon / 5' of anticodon) → Flexible "wobble" pairing
Source: Biochemistry, Lippincott Illustrated Reviews 8th ed, p. 1251-1253
Y not from vasudevan
Finding Sources
Finding Sources
Unfortunately, Vasudevan's Textbook of Biochemistry is not available in the library. The biochemistry books currently in the collection are:
| Book | Authors |
|---|---|
| Biochemistry, 8th ed (Lippincott Illustrated Reviews) | Denise Ferrier |
| Harper's Illustrated Biochemistry, 32nd ed | Murray, Rodwell et al. |
| Basic Medical Biochemistry - A Clinical Approach, 6e | Lieberman & Peet |
Vasudevan (full title: Textbook of Biochemistry for Medical Students by DM Vasudevan, Sreekumari & Kannan) is a very popular book especially for Indian medical students, but it hasn't been added to this library yet.
For now, I can answer your biochemistry questions from Lippincott or Harper's, which are equally comprehensive and widely used. If you'd like, I can cover the wobble hypothesis or any other topic from those sources!
Satyanarayana
Finding Sources
Satyanarayana's Biochemistry (by U. Satyanarayana & U. Chakrapani) is also not available in this library, unfortunately.
So the two most popular Indian biochemistry textbooks - Vasudevan and Satyanarayana - are both absent from the collection. The library currently has:
| Available Biochemistry Books |
|---|
| Lippincott Illustrated Reviews: Biochemistry (8th ed) |
| Harper's Illustrated Biochemistry (32nd ed) |
| Basic Medical Biochemistry - A Clinical Approach (6th ed) |
These are all excellent international references. The content - especially from Lippincott - covers all the same topics as Satyanarayana and Vasudevan, often in more detail with better diagrams.
Just ask your topic and I'll answer it from the available books!
This is a shared conversation. Sign in to Orris to start your own chat.