mechanism of contraction and relaxation of skeletal muscle.
Reading File
Finding Sources
Finding Sources
Reading File
Reading File
Finding Sources
Reading File
Reading File
Reading File
Loading Image
Loading Image
I now have comprehensive, detailed content from two authoritative physiology textbooks plus two excellent diagrams. Let me compose the full answer.
Mechanism of Contraction and Relaxation of Skeletal Muscle
1. Structural Basis: The Sarcomere
Each skeletal muscle fiber contains hundreds to thousands of myofibrils, each built from repeating units called sarcomeres - the functional units of contraction. Within each sarcomere:
- Thick filaments (myosin): ~1,500 per sarcomere, 1.6 µm long. Each myosin molecule has a double-helix tail and two globular heads (cross-bridges) that project outward. The heads contain an ATPase site and an actin-binding site.
- Thin filaments (actin): ~3,000 per sarcomere, attached to Z disks at each end. Actin is decorated with two regulatory proteins:
- Tropomyosin: a long protein that physically blocks the myosin-binding sites on actin at rest.
- Troponin complex (TnT, TnI, TnC): TnC is the Ca²⁺-sensing subunit; when Ca²⁺ binds, it shifts tropomyosin out of the blocking position.
- Titin filaments: elastic proteins that anchor myosin to the Z disk, providing passive elasticity and a series elastic element for force transmission.
A and I bands, H zones, and Z disks form the characteristic striated pattern visible on microscopy.
2. Excitation-Contraction (E-C) Coupling
This is the sequence linking a motor nerve signal to mechanical force. The steps always occur in strict temporal order: action potential → Ca²⁺ rise → contraction.
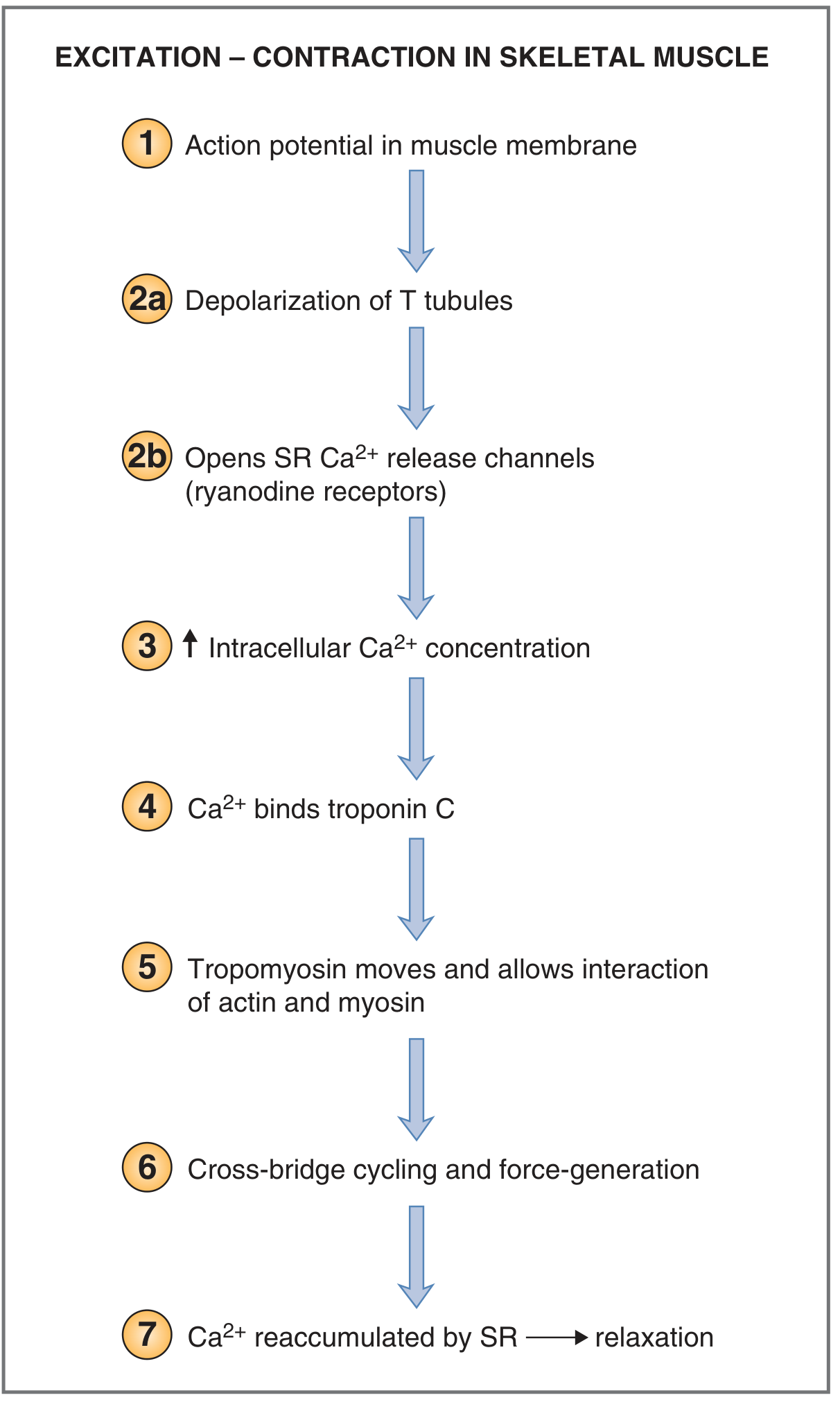
Step 1 - Motor Neuron Fires an Action Potential
The motor neuron releases acetylcholine (ACh) at the neuromuscular junction. ACh binds nicotinic receptors on the sarcolemma, generating an end-plate potential that triggers an action potential in the muscle fiber.
Step 2 - Propagation into T-Tubules
The action potential spreads along the sarcolemma and is carried deep into the fiber via the transverse (T) tubules - invaginations of the sarcolemma that run perpendicular to the myofibrils and contact the sarcoplasmic reticulum (SR) at structures called triads.
Step 3 - Voltage Sensor Activation (DHPR → RyR)
Depolarization of the T tubule membrane causes a conformational change in dihydropyridine receptors (DHPR) - voltage-sensing L-type Ca²⁺ channels. In skeletal muscle, the DHPR is physically coupled (via protein-protein contact) to ryanodine receptors (RyR1) on the SR membrane. The DHPR conformational change directly opens the RyR1 channels - no Ca²⁺ influx from the extracellular space is required (unlike cardiac muscle).
Step 4 - Ca²⁺ Release from the SR
RyR1 channels open, releasing Ca²⁺ from the terminal cisternae of the SR into the cytoplasm. Resting intracellular [Ca²⁺] is < 10⁻⁷ M; it rises to 10⁻⁷ - 10⁻⁶ M during activation.
Step 5 - Ca²⁺ Binds Troponin C
Ca²⁺ binds to troponin C (up to 4 Ca²⁺ ions per TnC molecule, cooperatively). This causes a conformational change in the troponin complex, which shifts tropomyosin away from the myosin-binding grooves on actin, exposing the active sites on actin.
3. The Cross-Bridge Cycle (Sliding Filament Mechanism)
With actin sites now exposed, the myosin heads undergo repeated attach-pull-detach cycles, each consuming one ATP. Actin filaments slide toward the center of the sarcomere (the "sliding filament theory"), shortening the sarcomere without any change in filament length.
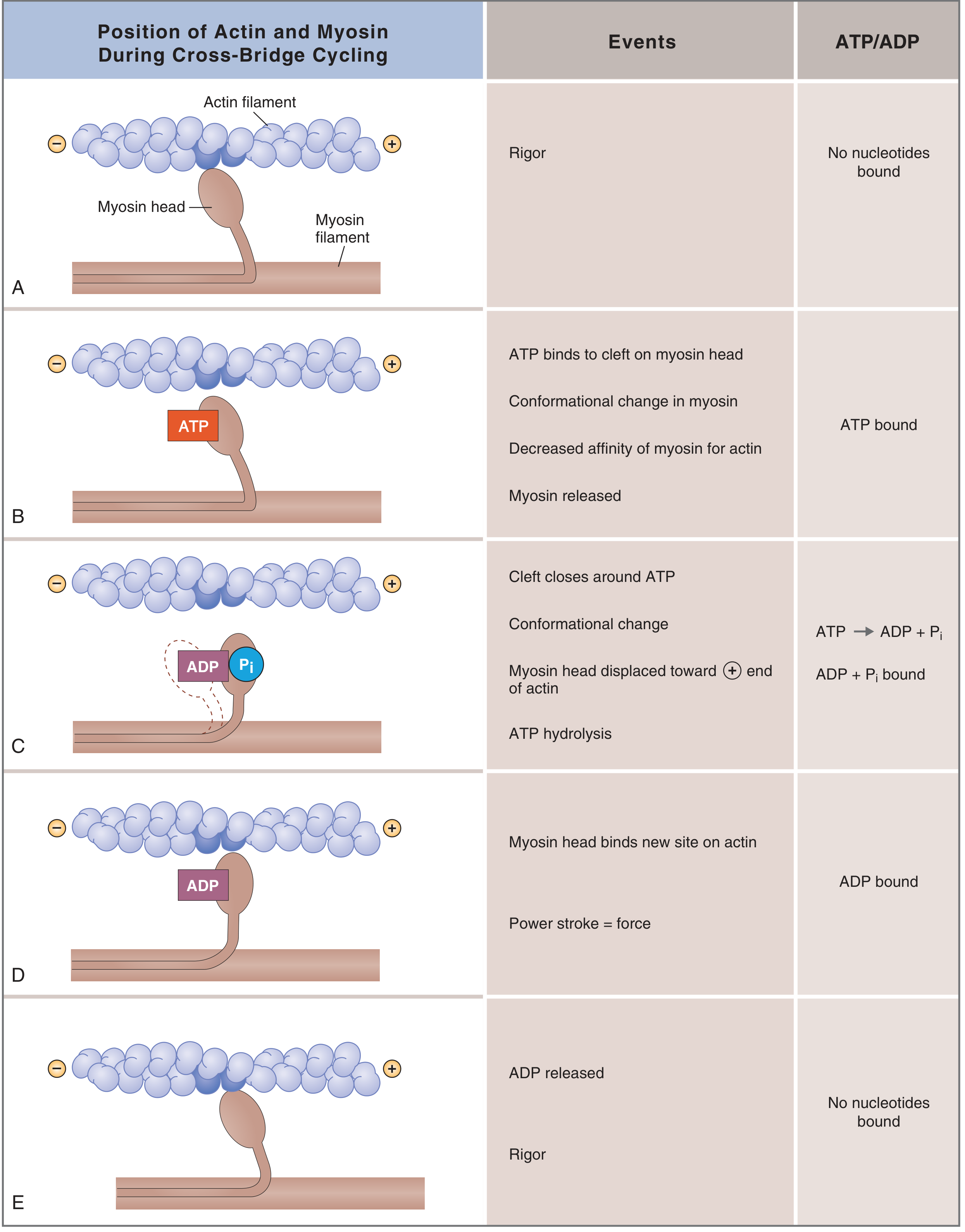
The five states of the cross-bridge cycle:
| State | Events | Nucleotide |
|---|---|---|
| A - Rigor | Myosin head tightly bound to actin; no ATP. This is the permanent state in rigor mortis. | None |
| B - Detachment | ATP binds to a cleft on the back of the myosin head → conformational change → decreased affinity for actin → myosin detaches from actin. | ATP bound |
| C - Cocking (repriming) | The cleft closes around ATP → ATP is hydrolyzed to ADP + Pi (both remain on myosin) → the myosin head rotates to a "cocked" (90°) high-energy position, displaced toward the plus end of the actin filament. | ADP + Pi bound |
| D - Power stroke | The cocked myosin head binds a new site on actin further toward the plus end → releases Pi → the head snaps from 90° to 45°, generating ~10 nm of movement and pulling the actin filament. This is the force-generating step. | ADP bound |
| E - ADP release → back to rigor | ADP is released → myosin returns to tight rigor state on actin, completing the cycle. | None |
Each cross-bridge cycle moves the actin filament ~10 nm and produces a force of ~1-4 pN per head. Because hundreds of cross-bridges cycle asynchronously, the cumulative force is smooth and sustained.
Key point on ATP: ATP is needed for two functions - (1) detaching myosin from actin (Step B), and (2) energizing the cocked position (Step C). Without ATP (death, ischemia), myosin remains locked to actin - rigor mortis.
4. Length-Tension Relationship
The force generated depends on the degree of actin-myosin filament overlap:
- Optimal length (~2.0-2.2 µm sarcomere length): maximal cross-bridge overlap → maximal tension.
- Overstretched (>2.2 µm): actin filaments pulled away from myosin heads → fewer cross-bridges engaged → tension falls.
- Overshortened (<1.65 µm): actin filaments from both sides begin to overlap each other, and Z disks abut the myosin filaments → tension falls sharply.
5. Relaxation
Relaxation is an active, ATP-dependent process:
- The motor neuron stops firing → ACh is no longer released at the NMJ.
- The muscle action potential ceases → T-tubules repolarize → DHPR returns to resting conformation → RyR1 channels close.
- SERCA (Sarco/Endoplasmic Reticulum Ca²⁺-ATPase) pumps Ca²⁺ back into the SR lumen against a concentration gradient, consuming ATP. Intracellular [Ca²⁺] falls below 10⁻⁷ M.
- Ca²⁺ dissociates from troponin C.
- Troponin I-mediated inhibition returns → tropomyosin shifts back over the myosin-binding sites on actin → active sites are blocked.
- Cross-bridge cycling stops → actin and myosin filaments passively slide back (assisted by titin's elasticity and external load) → sarcomere lengthens → muscle relaxes.
The series elastic elements (titin, connective tissue) must be stretched first for force to reach the surface, causing a brief delay between cross-bridge activity and observable tension - and a similar delay in tension fall after cycling stops.
Summary Table
| Phase | Key Event | Molecule Involved |
|---|---|---|
| Excitation | AP propagates into T-tubules | Sarcolemma, T-tubules |
| Coupling | DHPR conformational change → opens RyR1 | DHPR, RyR1 |
| Ca²⁺ release | Ca²⁺ floods cytoplasm from SR | SR terminal cisternae |
| Activation | Ca²⁺ binds TnC → tropomyosin shifts | Troponin C, tropomyosin |
| Contraction | Cross-bridge cycling (power stroke) | Myosin ATPase, actin |
| Relaxation | SERCA pumps Ca²⁺ back into SR | SERCA (ATP-dependent) |
Sources:
- Guyton and Hall Textbook of Medical Physiology, Chapter 6 (pp. 92-97)
- Costanzo Physiology 7th Edition, Chapter 1 (pp. 44-46)
This is a shared conversation. Sign in to Orris to start your own chat.