H
Hey! How can I help you today?
Can you access dm vasudevan textbook of biochemistry
Reading File
Finding Sources
Finding Sources
Unfortunately, DM Vasudevan's Textbook of Biochemistry is not available in the medical library. The library does have three biochemistry textbooks though:
| # | Title | Details |
|---|---|---|
| 1 | Basic Medical Biochemistry - A Clinical Approach, 6e | (Lieberman & Peet) |
| 2 | Biochemistry, 8th ed - Lippincott Illustrated Reviews | (popular Indian med school reference) |
| 3 | Harper's Illustrated Biochemistry, 32nd Ed | (classic standard text) |
Lippincott's Illustrated Reviews Biochemistry is probably the closest alternative to Vasudevan in terms of style and coverage for Indian medical students. Would you like me to look up a specific topic from any of these? I'm happy to search across all three for whatever you need.
Look up a specific biochemistry topic from available textbooks
Asking for Preferences
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Loading Image
Here is a thorough summary of DNA Replication & Repair compiled from the biochemistry textbooks in the library.
DNA Replication & Repair
Sources: Biochemistry, 8th ed - Lippincott Illustrated Reviews (p. 1135-1181) | Basic Medical Biochemistry - A Clinical Approach, 6e (p. 411-417)
The Central Dogma
The flow of genetic information follows the pattern: DNA → RNA → Protein. DNA replicates itself (replication), is copied into RNA (transcription), and RNA is decoded into protein (translation).
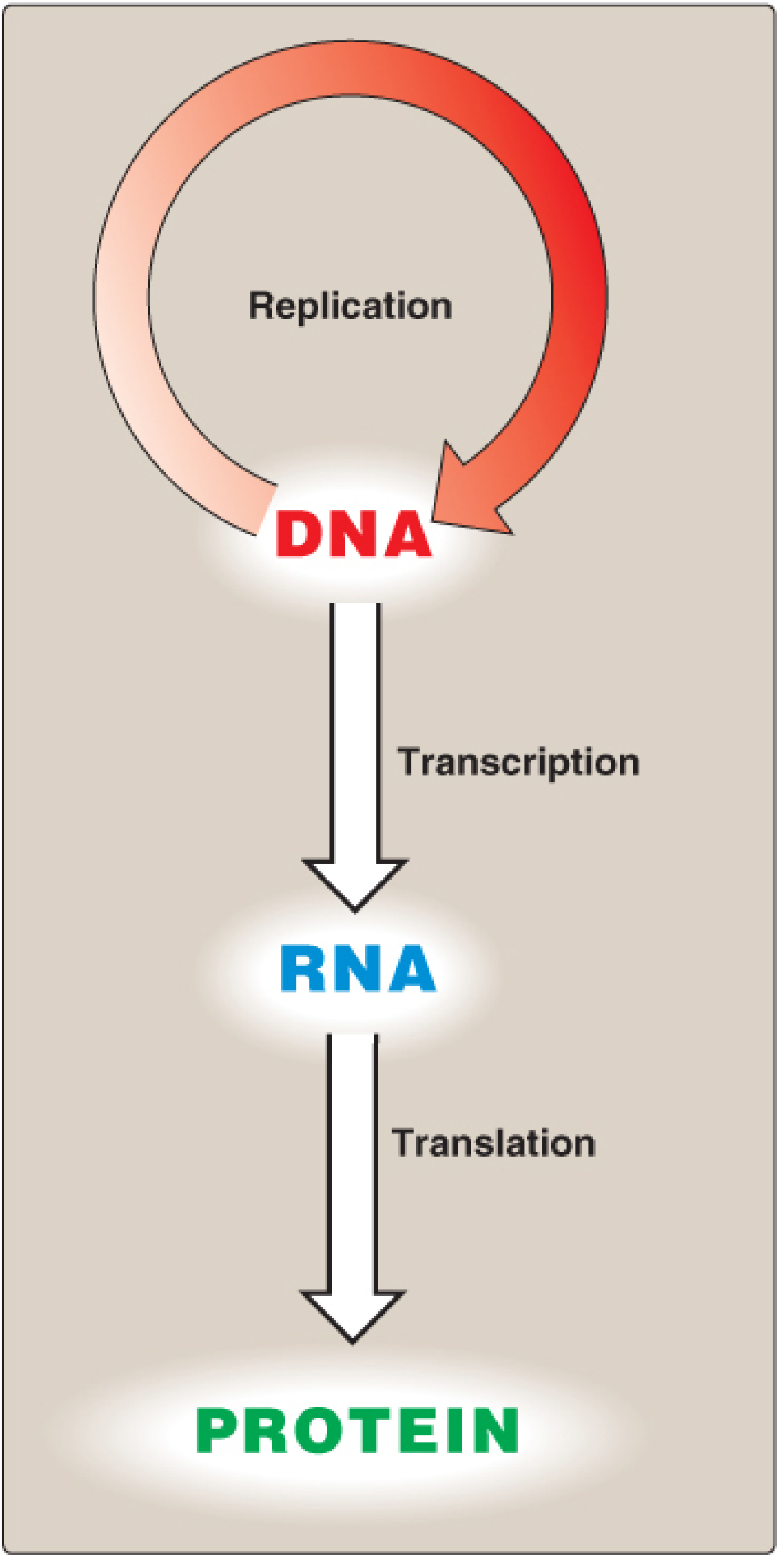
Part 1: DNA Replication
Key Concept: Semiconservative Replication
When two strands of double-stranded DNA separate, each acts as a template for a new complementary strand. The result is two daughter molecules, each containing one old (parental) strand and one newly synthesized strand. This is called semiconservative replication.
Steps in Prokaryotic DNA Replication (E. coli model)
1. Origin of Replication (oriC)
- Replication begins at a unique site called the origin of replication (ori)
- In prokaryotes: single origin (oriC); in eukaryotes: multiple origins per chromosome (allows faster replication of long DNA)
- The ori contains AT-rich segments that facilitate strand melting (AT pairs have only 2 hydrogen bonds vs. 3 in GC pairs)
2. Strand Separation
- DNA helicase unwinds and separates the two strands
- Single-strand binding proteins (SSBPs) stabilize the separated strands
- Topoisomerases relieve tension ahead of the replication fork
3. Replication Fork Formation
- Two replication forks form and move bidirectionally away from the origin, creating a replication bubble
- Each fork is a Y-shaped structure where synthesis is occurring
4. Primer Synthesis
- DNA polymerases cannot start a new chain; they can only extend an existing one
- Primase (a RNA polymerase) synthesizes a short RNA primer (~10 nucleotides) to provide a free 3'-OH for DNA pol to extend
5. Chain Elongation
- DNA Pol III (the main replicative polymerase in prokaryotes) synthesizes DNA 5' → 3' using each template strand
- Leading strand: synthesized continuously in the direction of fork movement
- Lagging strand: synthesized discontinuously as Okazaki fragments (short ~1000-2000 nt in prokaryotes, ~100-200 nt in eukaryotes) because it must be synthesized in the opposite direction
6. Primer Removal & Gap Filling
- DNA Pol I removes RNA primers with its 5'→3' exonuclease activity and fills in the gaps using its polymerase activity
- DNA Ligase seals the nicks between Okazaki fragments, joining the 3'-OH to the 5'-phosphate
7. Eukaryotic Replication
- Uses DNA Pol α (primase activity), DNA Pol δ (lagging strand), DNA Pol ε (leading strand)
- PCNA (proliferating cell nuclear antigen) acts as a sliding clamp, analogous to the β-clamp in prokaryotes
Proofreading
DNA Pol III has 3'→5' exonuclease (proofreading) activity - if a wrong base is incorporated, the enzyme detects the mismatch, excises the incorrect nucleotide, and inserts the correct one. This reduces errors to ~1 in 10⁷ nucleotides.
Part 2: DNA Repair
DNA sustains thousands of potentially mutagenic lesions per cell per day from:
- Chemicals: nitrous acid (deaminates bases), carcinogens like benzo[a]pyrene (forms bulky adducts with guanine)
- Radiation: UV light (pyrimidine dimers), ionizing radiation/X-rays (double-strand breaks, via hydroxyl radicals)
- Spontaneous changes: depurination, deamination
All excision repair systems share common steps:
- Recognize the DNA lesion
- Remove the damaged bases/nucleotides
- Resynthesise using the intact complementary strand as template (5'→3')
- Ligate to restore strand continuity
A. Mismatch Repair (MMR)
Corrects replication errors that escape proofreading (mismatched base pairs). Reduces error rate from 1 in 10⁷ to 1 in 10⁹ nucleotides.
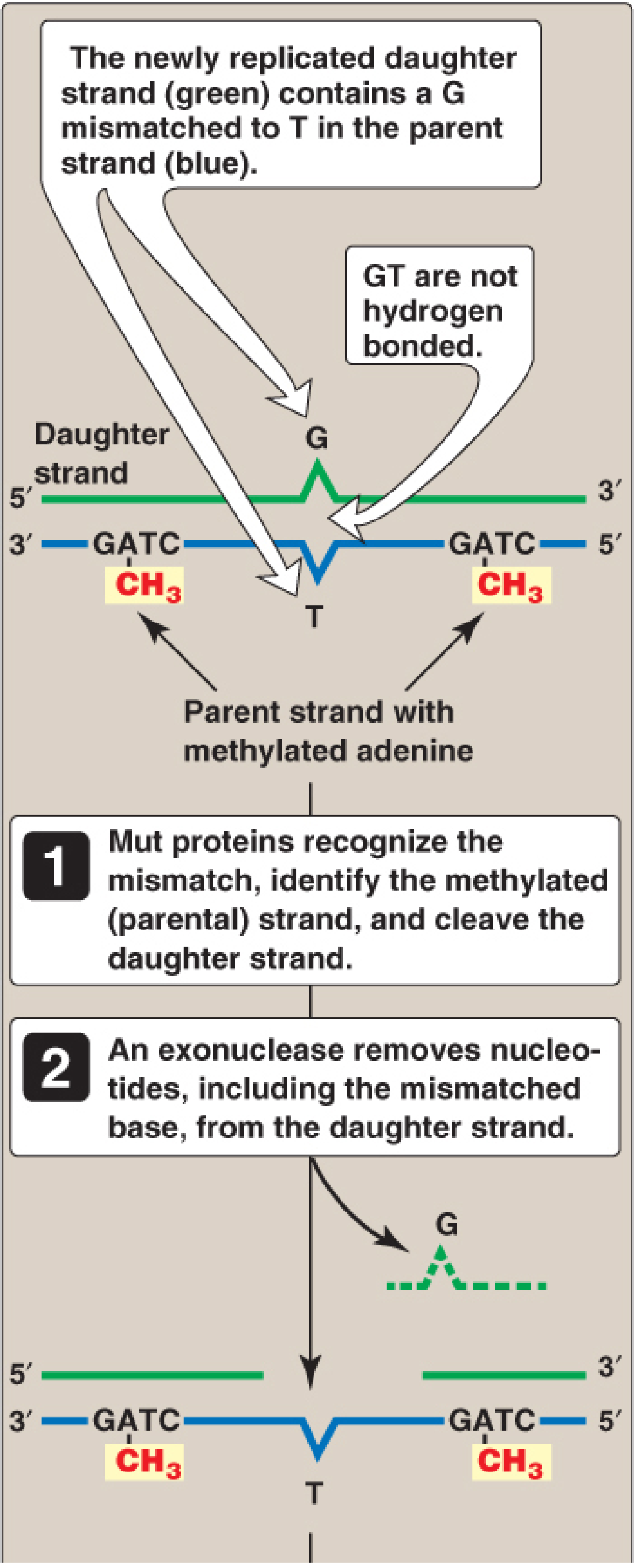
- Strand discrimination: In prokaryotes, GATC sequences on the parental strand are methylated (by DAM methylase) but the newly synthesized daughter strand is not yet methylated (hemimethylated). Mut proteins use this to identify which strand to repair.
- Mut proteins (MutS, MutL, MutH): MutS recognizes the mismatch, MutL recruits MutH, MutH cleaves the unmethylated daughter strand
- An exonuclease removes the mismatch; DNA Pol III fills the gap; DNA ligase seals it
Clinical link: Defective MMR genes in humans (MSH2, MLH1) cause Lynch syndrome (HNPCC) - hereditary nonpolyposis colorectal cancer. MSH2 and MLH1 mutations account for 90% of Lynch syndrome cases. About 5% of all colon cancer results from MMR mutations.
B. Nucleotide Excision Repair (NER)
Repairs bulky helix-distorting lesions such as:
- Thymine dimers (from UV radiation) - adjacent thymines on the same strand form covalent bonds
- Bulky adducts (e.g., benzo[a]pyrene-guanine adducts from cigarette smoke)
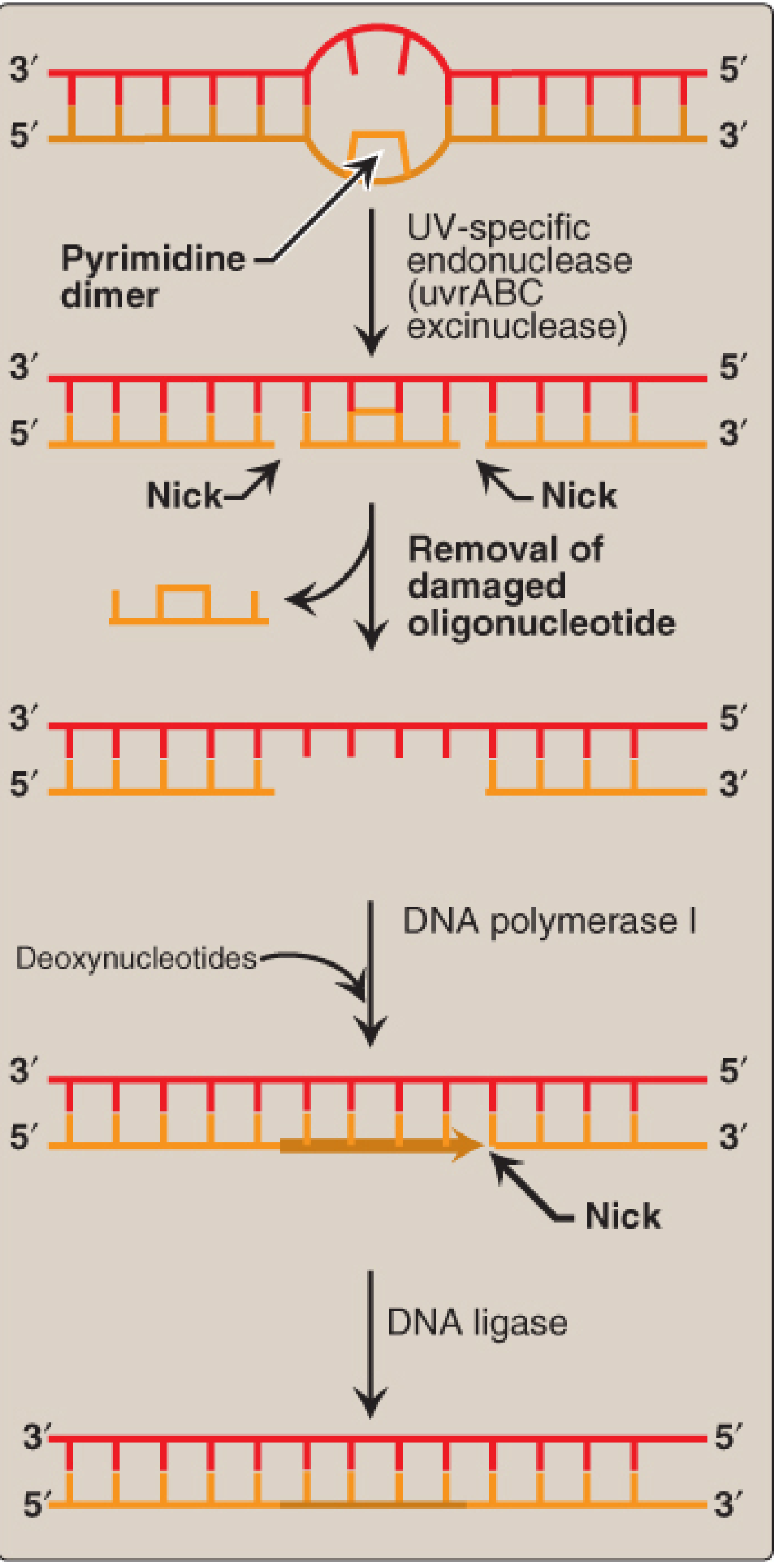
- In bacteria: UvrABC excinuclease (excinuclease complex) nicks the strand on both sides of the dimer, removing ~12 nucleotides. DNA Pol I fills the gap; DNA ligase seals it.
- In humans: XP proteins (XPA through XPG) perform an analogous function, removing ~25-30 nucleotides
- Clinical link: Defects in NER genes cause Xeroderma Pigmentosum (XP) - extreme UV sensitivity and high rates of skin cancer
C. Base Excision Repair (BER)
Removes and replaces individual damaged bases (oxidized, deaminated, or alkylated bases) without removing the entire nucleotide.
- A DNA glycosylase cleaves the glycosidic bond between the altered base and the sugar, leaving an apurinic/apyrimidinic (AP) site
- AP endonuclease nicks the backbone
- DNA pol fills the gap; ligase seals
D. Photoreactivation (Bacteria only)
Bacteria can reverse thymine dimers directly using photolyase enzymes that use visible light energy to cleave the abnormal covalent bonds between pyrimidines. No nucleotides are removed - it is a direct reversal, not an excision mechanism.
E. Double-Strand Break Repair
For double-strand breaks (from ionizing radiation):
- Homologous recombination (HR): uses a homologous sequence as template; accurate; occurs in S and G2 phases
- Non-homologous end joining (NHEJ): directly joins broken ends; error-prone; occurs throughout the cell cycle
Summary Table: DNA Repair Mechanisms
| Mechanism | Lesion Repaired | Key Proteins | Clinical Disease if Defective |
|---|---|---|---|
| Mismatch Repair | Replication mismatches | MutS/MutL/MutH (E. coli); MSH2/MLH1 (humans) | Lynch syndrome (HNPCC) |
| Nucleotide Excision Repair | Bulky adducts, thymine dimers | UvrABC (E. coli); XP proteins (humans) | Xeroderma Pigmentosum |
| Base Excision Repair | Oxidized/deaminated bases | DNA glycosylase, AP endonuclease | - |
| Photoreactivation | Thymine dimers | Photolyase | Bacteria only |
| Homologous Recombination | Double-strand breaks | BRCA1/2, RAD51 | Breast/ovarian cancer risk |
| Non-Homologous End Joining | Double-strand breaks | Ku proteins, DNA-PK | - |
- Biochemistry 8th ed Lippincott Illustrated Reviews, p. 1135-1181
- Basic Medical Biochemistry - A Clinical Approach 6e, p. 411-417
Would you like me to go deeper on any specific aspect - for example, the specific DNA polymerases, telomere replication, or a particular repair disorder?
This is a shared conversation. Sign in to Orris to start your own chat.