Haemoglobin and respiration
Reading File
Finding Sources
Finding Sources
Searching the Web
haemoglobin structure function oxygen transport respiration physiology
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Finding Sources
Reading File
Reading File
Reading File
Reading File
Reading File
Loading Image
Loading Image
Reading File
Loading Image
Reading File
I now have rich content from multiple authoritative textbooks. Let me compile the comprehensive answer.
Haemoglobin and Respiration
1. Structure of Haemoglobin
Haemoglobin (Hb) is a tetrameric globular protein found exclusively in red blood cells. The adult form, HbA, consists of two α-chains and two β-chains (α₂β₂), each holding a heme prosthetic group - making four O₂ binding sites per molecule.
The Heme Group
Each heme unit is an iron-porphyrin compound: a planar ring of four pyrrole groups linked by methyne bridges, with a central Fe²⁺ ion. This deep-red chromophore gives blood its colour.
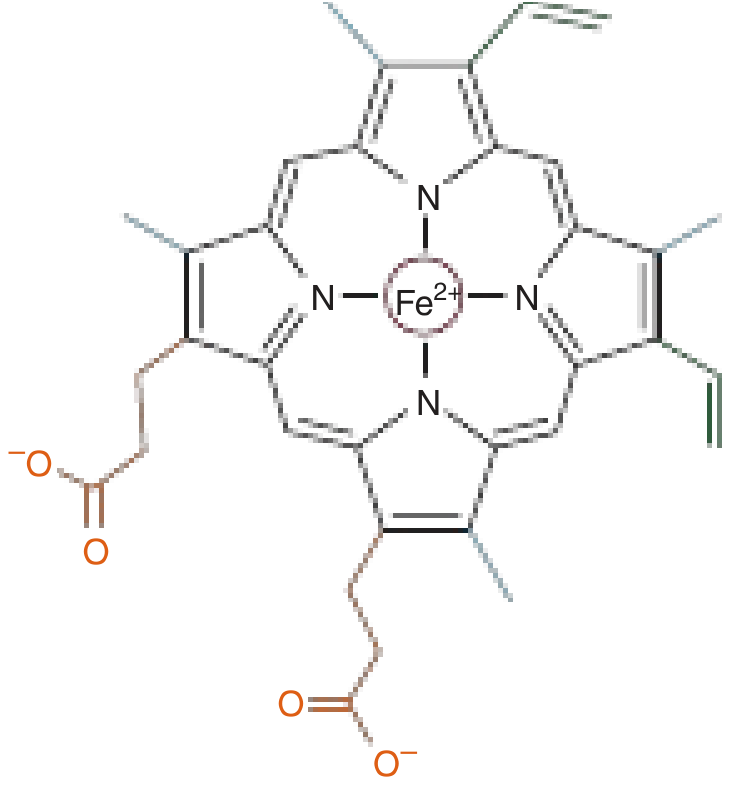
- The iron must remain in the ferrous (Fe²⁺) state to bind O₂. Oxidation to Fe³⁺ (forming methaemoglobin) abolishes oxygen-carrying capacity.
- The fifth coordination position of iron is bonded to the proximal histidine (His F8) of the globin chain.
- The distal histidine (His E7) lies on the opposite side and prevents CO₂ from irreversibly oxidising Fe²⁺ to Fe³⁺.
(Harper's Illustrated Biochemistry, 32nd Ed.)
T and R States (Cooperativity)
The haemoglobin tetramer exists in two conformations:
| State | Name | O₂ Affinity | Structure |
|---|---|---|---|
| T (Tense/Taut) | Deoxy-Hb | Low | Fe²⁺ pulled out of heme plane; ionic + H-bonds between αβ dimers |
| R (Relaxed) | Oxy-Hb | High | Fe²⁺ pulled into heme plane; polar bonds between dimers broken |
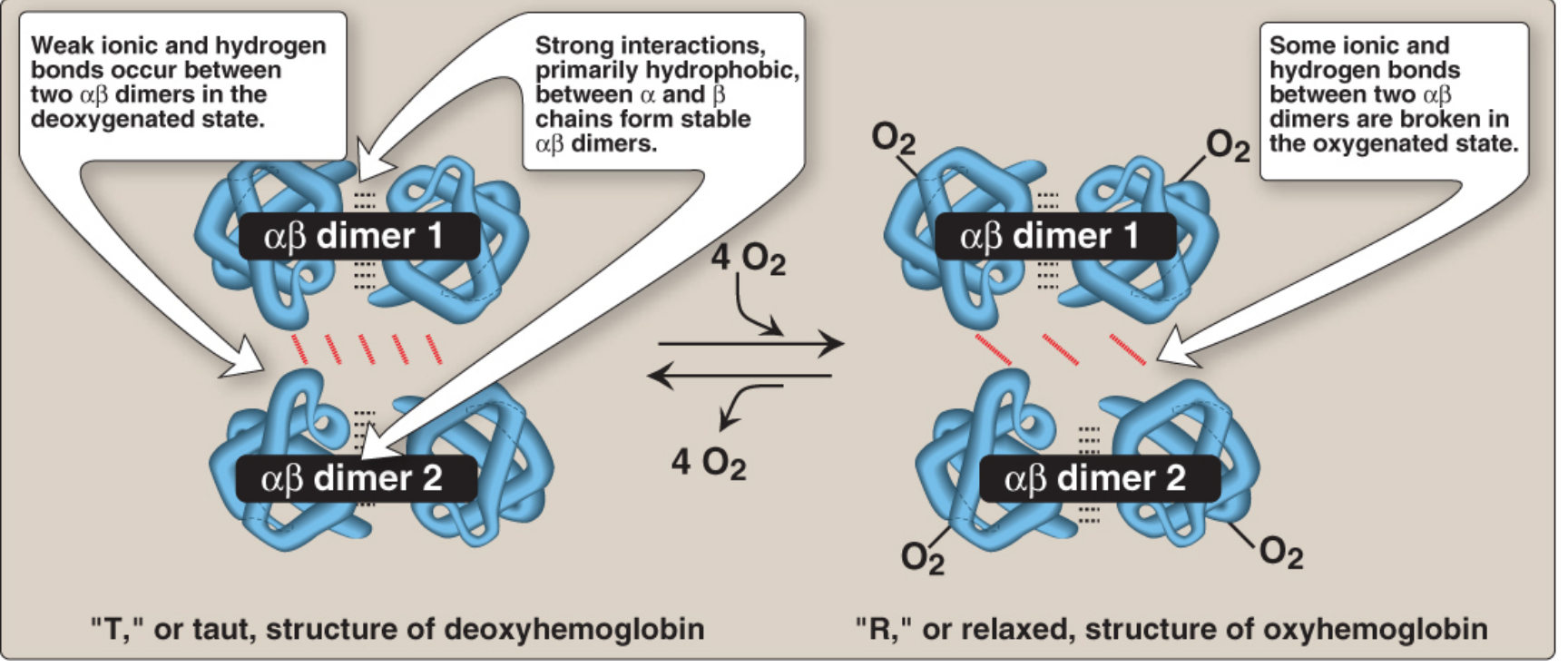
When the first O₂ binds, the Fe²⁺ pulls into the heme plane, moving the proximal histidine and triggering a conformational shift in adjacent subunits - increasing their O₂ affinity. This is positive cooperativity, producing the characteristic sigmoidal oxygen-dissociation curve (ODC).
(Lippincott's Biochemistry, 8th Ed.)
2. The Oxygen-Dissociation Curve (ODC)
Oxygen is carried in blood in two ways:
- Bound to haemoglobin (by far the dominant form - ~98%)
- Dissolved in plasma (very small amount, proportional to PO₂ by Henry's law)
The ODC plots % saturation of Hb with O₂ against partial pressure of O₂ (PO₂). Its S-shape reflects cooperative binding:
- At alveolar PO₂ ~100 mmHg: Hb is ~97-98% saturated (efficient O₂ loading)
- At tissue/venous PO₂ ~40 mmHg: Hb drops to ~75% saturated (efficient O₂ unloading)
- P₅₀ = PO₂ at which Hb is 50% saturated; normal ~26-27 mmHg in adults
Factors Shifting the ODC
Right shift = decreased O₂ affinity = more O₂ released to tissues (higher P₅₀)
| Factor | Direction | Physiological Meaning |
|---|---|---|
| ↑ PCO₂ | Right | Active tissues release more O₂ |
| ↓ pH (acidosis) | Right | Bohr effect (see below) |
| ↑ Temperature | Right | Exercising muscle gets more O₂ |
| ↑ 2,3-BPG | Right | High altitude adaptation |
| Left shift | ↑ affinity | Less O₂ released |
| ↓ PCO₂, ↑ pH | Left | In lungs - favours O₂ loading |
| Fetal Hb (HbF) | Left | P₅₀ = 18 mmHg (vs 27 mmHg adult) |
| HbS (sickle) | Right | Reduced affinity |
(Murray & Nadel's Textbook of Respiratory Medicine; Lippincott's Biochemistry)
3. The Bohr Effect
In metabolically active tissues, CO₂ production rises and pH falls. This causes a right shift of the ODC, promoting O₂ unloading exactly where it is needed most.
Mechanism:
- CO₂ + H₂O → H₂CO₃ → HCO₃⁻ + H⁺ (catalysed by carbonic anhydrase in RBCs)
- H⁺ binds to specific histidine residues (higher pKa in deoxyHb), forming ionic salt bridges that stabilise the T (deoxy) state
- This reduces O₂ affinity and promotes O₂ release
The equation:
HbO₂ + H⁺ ⇌ HbH⁺ + O₂
HbO₂ + H⁺ ⇌ HbH⁺ + O₂
Conversely, in the lungs, CO₂ is exhaled, pH rises, and the curve shifts left, favoring O₂ loading.
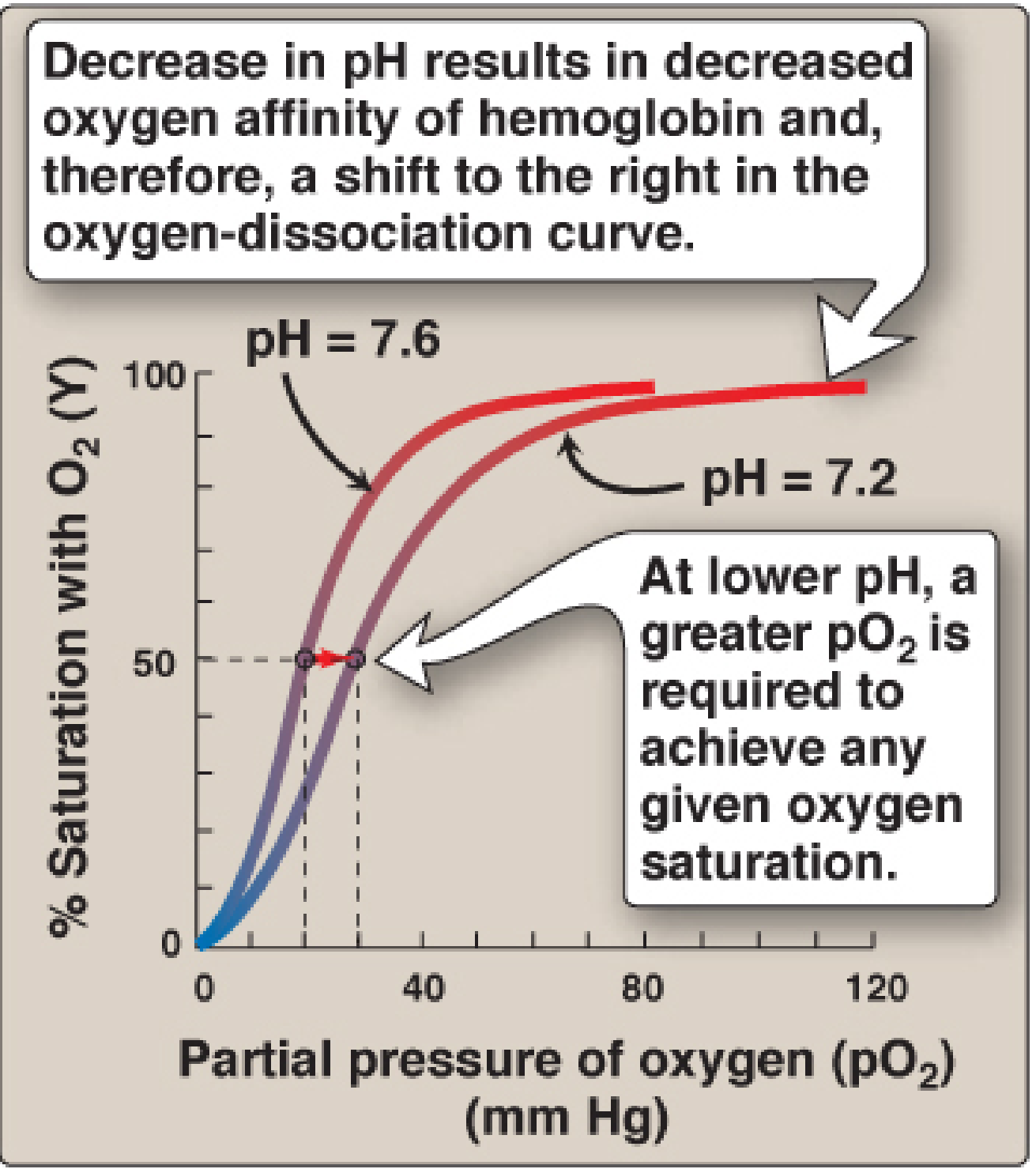
(Lippincott's Biochemistry, 8th Ed.)
4. Role of 2,3-Bisphosphoglycerate (2,3-BPG)
2,3-BPG is synthesised from a glycolytic intermediate in RBCs and is the most abundant organic phosphate in those cells (approximately equimolar with Hb).
- 2,3-BPG binds preferentially to the central cavity of deoxy-Hb (T state), forming ionic bonds with positively-charged residues of the β-chains
- This stabilises the T state, shifting the ODC right and reducing O₂ affinity
- At high altitude: hypoxia stimulates increased 2,3-BPG, right-shifting the ODC to enhance O₂ delivery to tissues
- Fetal Hb (HbF): has γ-chains instead of β-chains; γ-chains bind 2,3-BPG poorly, so HbF has higher O₂ affinity - ideal for extracting O₂ from maternal blood
(Lippincott's Biochemistry, 8th Ed.)
5. CO₂ Transport and the Haldane Effect
CO₂ is transported from tissues to lungs in three forms:
| Form | % of Total |
|---|---|
| As HCO₃⁻ (bicarbonate in plasma) | ~70% |
| Carbaminohaemoglobin (CO₂ bound to N-terminal amino groups of Hb) | ~20-25% |
| Dissolved in plasma | ~5-10% |
The Haldane Effect is the complement of the Bohr effect:
- Deoxygenated Hb binds CO₂ and H⁺ more avidly than oxygenated Hb
- At the tissues: Hb unloads O₂ → becomes better CO₂ carrier
- At the lungs: Hb loads O₂ → releases CO₂ and H⁺ for exhalation
This means O₂ loading in the lungs directly promotes CO₂ unloading - a beautifully coordinated reciprocal exchange.
(Murray & Nadel's Textbook of Respiratory Medicine)
6. Oxygen Cascade: Lungs to Mitochondria
The PO₂ falls progressively from air to the site of use:
| Location | PO₂ (mmHg) |
|---|---|
| Dry room air | ~159 |
| Humidified tracheal air | ~149 |
| Alveolar gas | ~100 |
| Arterial blood | ~95-100 |
| Mixed venous blood | ~40 |
| Mitochondria | <10 |
In a normal lung, blood transiting the pulmonary capillary (~0.75 seconds at rest) is fully oxygenated in the first one-third of capillary transit, providing substantial diffusion reserve. Under exercise or disease (thickened blood-gas barrier), this reserve may be exhausted and diffusion limitation can occur.
(Murray & Nadel's Textbook of Respiratory Medicine)
7. Variant Haemoglobins of Clinical Importance
| Haemoglobin | Feature | Clinical Significance |
|---|---|---|
| HbF (fetal) | γ-chains, low 2,3-BPG binding | High O₂ affinity; switches to HbA by ~6 months |
| HbS (sickle) | Glu→Val substitution β6 | Polymerises when deoxygenated → sickling, vaso-occlusion |
| HbA₂ | δ-chains | Elevated in β-thalassaemia trait |
| MetHb | Fe³⁺ (oxidised) | Cannot carry O₂; causes cyanosis |
| COHb | CO bound to Fe²⁺ | 250× affinity for CO vs O₂; left-shifts ODC (CO poisoning) |
(Murray & Nadel's; Tietz Laboratory Medicine, 7th Ed.)
Summary
Haemoglobin is the master molecule of respiratory gas transport. Its sigmoidal ODC - generated by cooperative binding across its four subunits - allows efficient O₂ loading in the lungs (high PO₂) and efficient O₂ unloading at active tissues (low PO₂, low pH, high CO₂, high temperature). The Bohr effect (pH/CO₂ shifting the ODC right) and Haldane effect (deoxygenation enhancing CO₂ carriage) are reciprocal mechanisms ensuring that wherever metabolism is highest, O₂ delivery is greatest and CO₂ removal is most efficient. 2,3-BPG provides an additional regulatory handle, particularly important at altitude or in chronic anaemia.
This is a shared conversation. Sign in to Orris to start your own chat.